Keywords
Aldose reductase, Protein unfolding, Folding intermediate, Cooperativity, Tryptophan fluorescence, ANS fluorescence, Thermal unfolding
Aldose reductase, Protein unfolding, Folding intermediate, Cooperativity, Tryptophan fluorescence, ANS fluorescence, Thermal unfolding
GuHCl, guanidine hydrochloride; TCEP, (tris(2-carboxyethyl)phosphine); ANS, 8-anilino-1-naphthalenesulfonic acid ammonium salt; IPTG, isopropyl β-D-1-thiogalactopyranoside; Trp, tryptophan.
Human aldose reductase (hAR) (EC 1.1.1.21) is an NADPH-dependent oxidoreductase that belongs to super family of aldo-keto reductases1. Being the first and rate limiting enzyme of polyol pathway, hAR converts glucose to sorbitol2. Under hyperglycemic conditions, the polyol pathway is up-regulated and a significant proportion of glucose gets fluxed through this pathway, which leads to accumulation of sorbitol, consumption of NADPH and redox imbalance of NADPH/NADP+ ratio. All these factors have been linked with various tissue based pathologies associated with secondary complications of diabetes mellitus3. Due to its clinical importance, hAR has been widely studied from the perspective of development of potent inhibitors so as to prevent or delay the onset of secondary diabetic complications4.
Extensive information is available in the literature about the structure and function of hAR, particularly related to active site of hAR from ultra-high-resolution crystal structures with a number of potential inhibitors5, flexibility in the hAR binding site pocket6 and the thermodynamics of closing/opening of the specificity pocket within binding site pocket of hAR7. Nevertheless, there is little investigation related to the folding/unfolding mechanism of hAR. Understanding the capability of a polypeptide chain to spontaneously fold into a compact tertiary structure on biological relevant time scale is a long-standing challenge in protein science8.
Under physiological conditions, protein structure fluctuates among different native conformations separated by close free energy barriers9. Since hAR activity leads to sorbitol accumulation, leading to osmotic stress; it seems to function under stress conditions which might perturb its native conformation ensemble. Here we report on thermally and chemically induced unfolding studies of hAR. Thermal unfolding revealed simple two-state transition whereas chemical induced unfolding led us to discover an intermediate state during hAR unfolding.
The hAR cDNA cloned into expression vector pET-15b (Novagen) was a kind gift from Dr. Alberto Podjarny (Department of Integrated Structural Biology, Institut de Génétique et de Biologie Moléculaire et Cellulaire, CNRS, INSERM, France). The plasmid, coding for a hexahistidine-tagged protein, was expressed into E. coli strain BL21 (DE3) (Novagen). The cells with recombinant plasmid were grown with 100 μM ampicillin at 37°C to an OD600 nm value of 0.7 and protein expression was induced by adding 1 mM IPTG. Cells were grown for further 3 hours at 37°C. All further operations were carried out at 4°C unless otherwise stated. Cells were centrifuged, re-suspended and lysed by sonication. A Ni-NTA affinity column (GE Healthcare) was used for protein purification. The material used for stationary phase for the column was Ni-Sepharose and the flow rate of column was adjusted to 0.5 ml min-1. Imidazole and other salts were removed by repeated dialysis in 50 mM potassium phosphate, pH-7 buffer containing 50 mM NaCl. Protein concentration was estimated using the molar extinction coefficient and absorbance reading at 280 nm. The histidine tag from recombinant protein was removed by thrombin (4 units of thrombin per mg of recombinant protein at room temperature for 3 hours). Cleaved protein was passed through the Ni-NTA column, and purified protein without tag was collected as flow-through. Enzyme activity was checked as per standard assay10. Homogeneity and molecular weight of hAR with and without histidine tags was analyzed under denaturing conditions on 15% SDS-PAGE. Purified hAR was stored at -20°C for further studies.
Thermal unfolding was carried out at a final concentration of 2.8 µM protein in 50 mM potassium phosphate buffer, pH 7.0 containing 50 mM NaCl and 0.1mM TCEP. Transition between 20–70 °C was followed using a far-UV circular dichroism (CD) signal at 222 nm by using 0.1 cm path length cuvette at sampling rate of 1.0 °C min-1 in a Jasco J-810 spectropolarimeter. Buffer blank was duly subtracted before reporting the change in ellipticity (millidegree) at 222 nm.
Samples with 1.4 μM protein concentration were prepared in phosphate buffer (described earlier) containing different concentrations of GuHCl/urea. Samples were incubated for 12 hours to reach equilibrium at 25°C after which no change in signal occurred either in fluorescence or CD spectra. Trp fluorescence (excitation at 295 nm and emission recorded between 300 nm to 400 nm) and ANS fluorescence (excitation at 370 nm and emission recorded between 400 nm to 600 nm) measurements were performed using a Hitachi F-7000 fluorescence spectrophotometer for GuHCl samples and Jasco J-815 spectropolarimeter for urea samples. Far-UV CD measurements were performed using Jasco J-810 spectropolarimeter for GuHCl samples and Jasco J-815 spectropolarimeter for urea samples. Quartz cuvette of 1 cm and 0.5 cm path length were used for fluorescence and CD measurements respectively. All measurements were done at 25 °C. Spectra were reported as ellipticity (millidegree) after baseline correction.
ΔCp value for unfolding of hAR was calculated from change in accessible surface area (ΔASA) according to Equation 111
ProtSA web server was used to calculate change in accessible surface area from native to unfolded conformation of hAR12.
GraphPad Prism version 7.04 for Windows (GraphPad Software, La Jolla, California) was used for analysis of thermal and chemical induced unfolding data on the basis of two and three state model respectively as described in following sections.
Data was fitted by least square analysis to Equation 213.
Where An and Au are native and unfolded state baseline intercepts respectively and bN and bU are native and unfolded baseline slopes respectively. ΔHm is enthalpy change at melting temperature (Tg). T is absolute temperature and R is the gas constant.
Signal for native (YN = An + bN × T) and unfolded baseline (YU = Au + bU × T) for every point in transition region was calculated from Equation 2. If Y is signal for a particular point in transition region, then fraction of unfolded protein (Fu) at this point is given by Equation 3.
The equilibrium constant (k) can be calculated from relative population of species using Equation 4.
ΔG can be calculated as a function of temperature using Equation 5.
The thermal stability curve of hAR was constructed on the basis of Equation 6–814.
Where ΔHT and ΔST are enthalpy and entropy change respectively at temperature T with reference to Tg. Th, Ts and Tg are the temperatures at which ∆H, ∆S, and ∆G are zero respectively. ∆Gs is the stabilization free energy of the native state relative to the unfolded state.
Data was fitted by least square analysis to Equation 913.
Where An, Au, and Ai are the native, unfolded and intermediate baseline intercepts, respectively, and bN, bU and bI are the native, unfolded and intermediate baseline slopes, respectively. [D] is denaturant concentration in molar. m(N-I) and m(I-U) are denaturant gradient for native to intermediate and intermediate to unfolded state respectively. ∆G(N-I) and ∆G(I-U) are stabilization free energy of intermediate state relative to native and unfolded state respectively.
Change in ellipticity at 222 nm fitted well on the basis of a two-state model (Figure 1A). This analysis gave values for ΔHg and Tg which along with ΔCp value calculated from Equation 1 were used for non-linear regression of transition region (±5 kJ mol-1) to Equation 8 (Figure 1C). Values of ΔH and ΔS were calculated over extended range of temperature by using Equation 7 and Equation 8, respectively (Figure 1D). Thermal stability curve is extrapolation of transition region assuming constant ΔCp during unfolding transition (Figure 1E). The relationship between Ts, Th and ∆G (Ts – Th = ∆Gs/∆Cp) is presented in Figure 1F. Thermodynamic parameters obtained from analysis of thermal unfolding data are listed in Table 1. All raw data are available as Underlying data15.
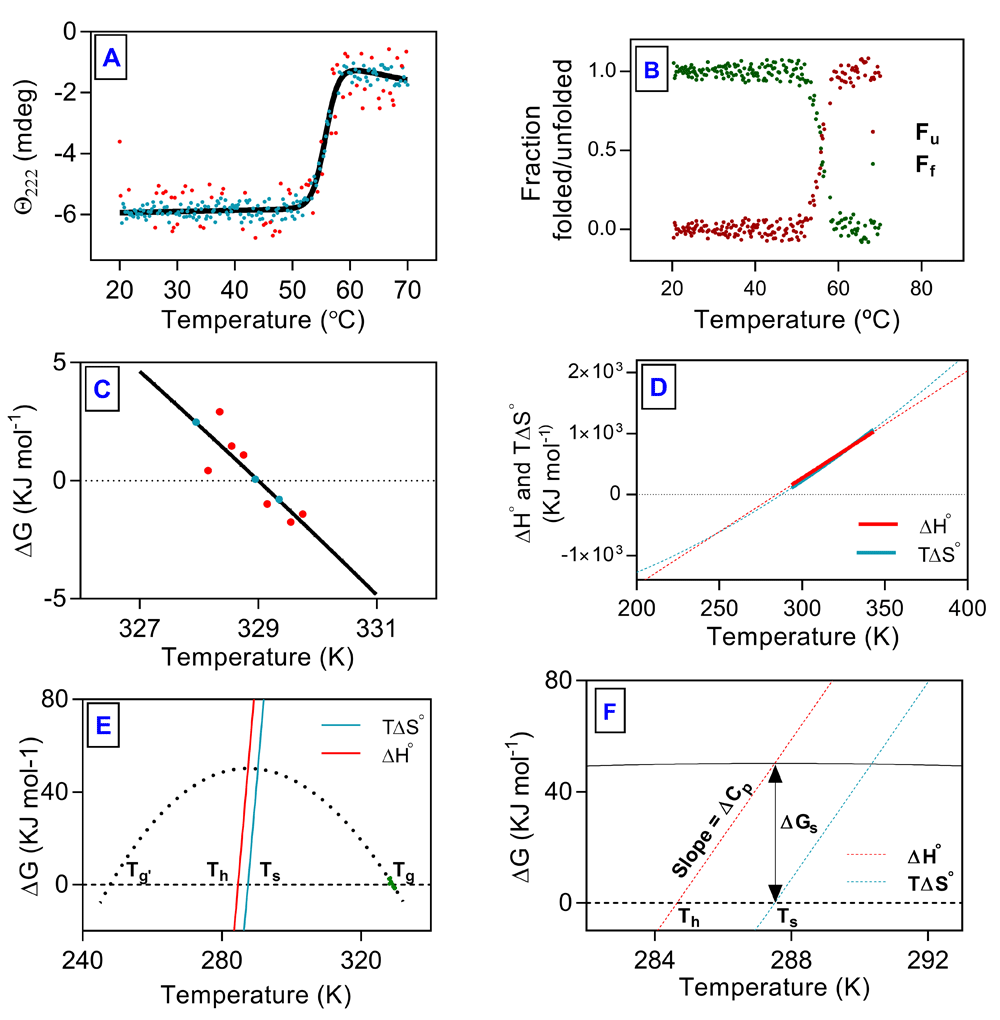
(A) Change in ellipticity at 222 nm plotted as a function of temperature. (B) Fraction of protein folded (green dots) and unfolded (red dots) plotted against temperature. (C) Portion of transition curve used in van't Hoff analysis. (D) Plots of ΔH and ΔS as function of temperature. (E) thermal stability curve of hAR. (F) Triangular relationship among Th, Ts and ∆Gs. Explanation for Tg, Tg’, Th and Ts is given in text. Dashed lines are extrapolations. Solid line represents fit to unfolding transition, filled symbols represent data points from unfolding experiments and red symbols represent outliers.
There are six Trp residues in hAR, out of which four are part of the hydrophobic active site pocket in the core of the β-barrel and two are buried in alpha helices surrounding the barrel. Their fluorescence provided global signal of change in tertiary structure. ANS has been extensively used as a probe for non-native, partially unfolded conformations of protein. The binding of ANS to hydrophobic regions results in a significant enhancement of ANS fluorescence and a pronounced blue-shift of the λmax16.
Fluorescence emission profiles of hAR equilibrated with different concentrations of denaturants are presented in Figure 2 (Figure 2A and Figure 3A for urea and GuHCl, respectively). A plot of λmax against denaturant concentration indicated cooperative transition from native to unfolded state (Figure 2B and Figure 3B for urea and GuHCl, respectively). In case of ANS fluorescence, significant blue-shift of around 20 nm and 10 nm from the native to intermediate state was observed for urea and GuHCl, respectively (Figure 2B2 and Figure 3B2 for urea and GuHCl, respectively).
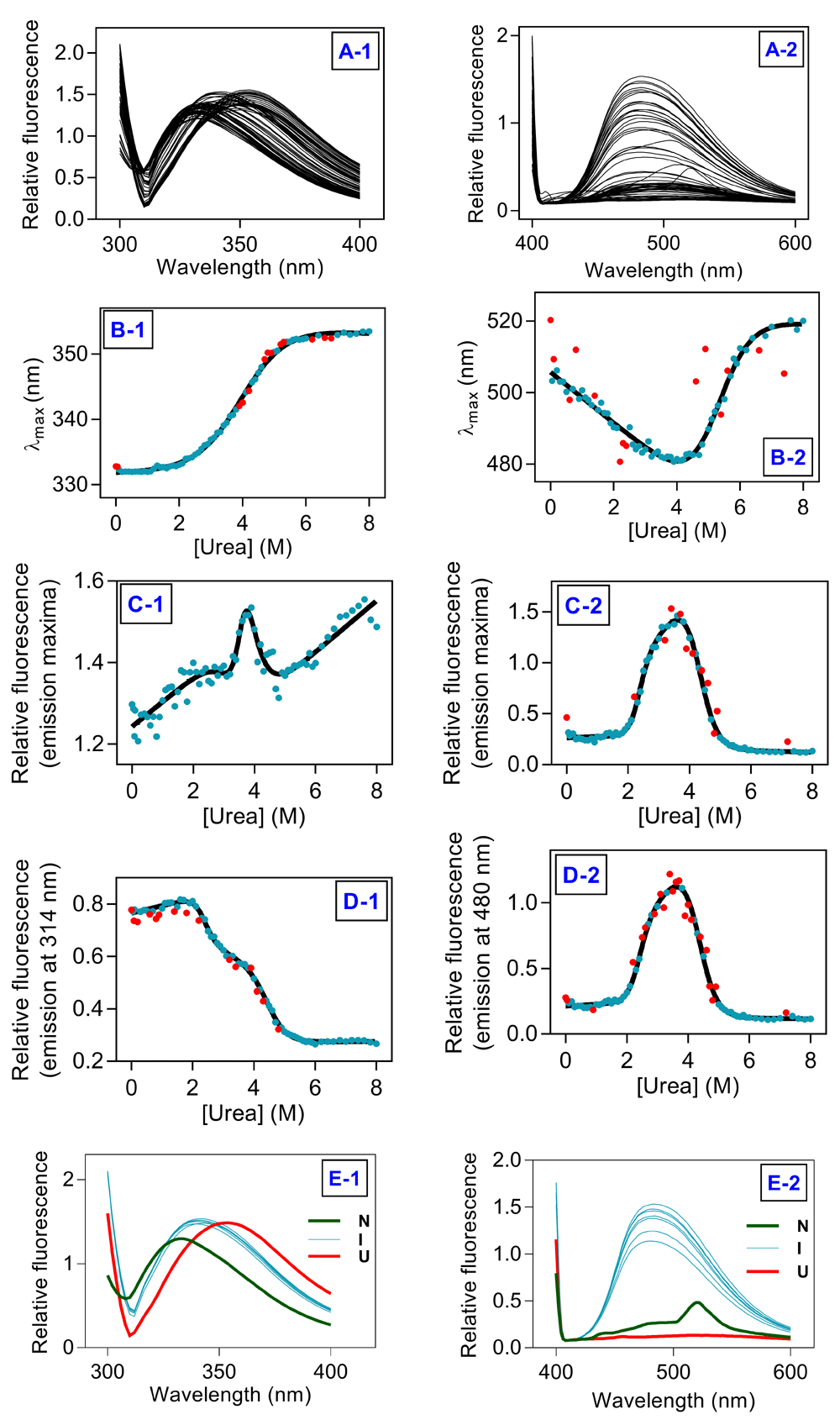
(A1) Trp fluorescence scans and (A2) ANS fluorescence scans for all the samples. (B1) λmax (Trp fluorescene) and (B2) λmax (ANS fluorescence) against [urea]. (C1) Imax (Trp fluorescenc) and (C2) Imax (ANS fluorescence) against [urea]. (D1) I295/314 (Trp fluorescence) and (D2) I370/480 (ANS fluorescence) against [urea]. (E1) Trp fluorescence and (E2) ANS fluorescence of samples in native (green), intermediate (cyan) and unfolded state (red). Solid lines represent fit to the unfolding transitions, filled symbols represent data points from unfolding experiments, red symbols represent outliers.
Plot of Imax against denaturant concentration indicated presence of an intermediate during unfolding transition (Figure 2C and Figure 3C for urea and GuHCl, respectively). Imax in case of ANS fluorescence fits satisfactorily on the basis of three-state model (Figure 2C2 and Figure 3C2 for urea and GuHCl, respectively). For both urea and GuHCl induced unfolding, Trp fluorescence emission intensity at 314 nm fits satisfactorily to three-state model. In case of ANS fluorescence, both Imax and fluorescence emission intensity at 480 nm fit equally well on the basis of three-state model. Thus, Trp fluorescence emission intensity at 314 nm and ANS fluorescence emission intensity at 480 nm were analyzed on the basis of three-state model to evaluate thermodynamic stability of hAR (Figure 2D and Figure 3D for urea and GuHCl, respectively). The thermodynamic parameters obtained from fitting are listed in Table 1. Trp and ANS fluorescence clearly demonstrate presence of an intermediate state populated at 3.5-4.0 M and 0.7-2 M urea and GuHCl concentration respectively, apart from the native and unfolded states (Figure 2E and Figure 3E for urea and GuHCl, respectively).
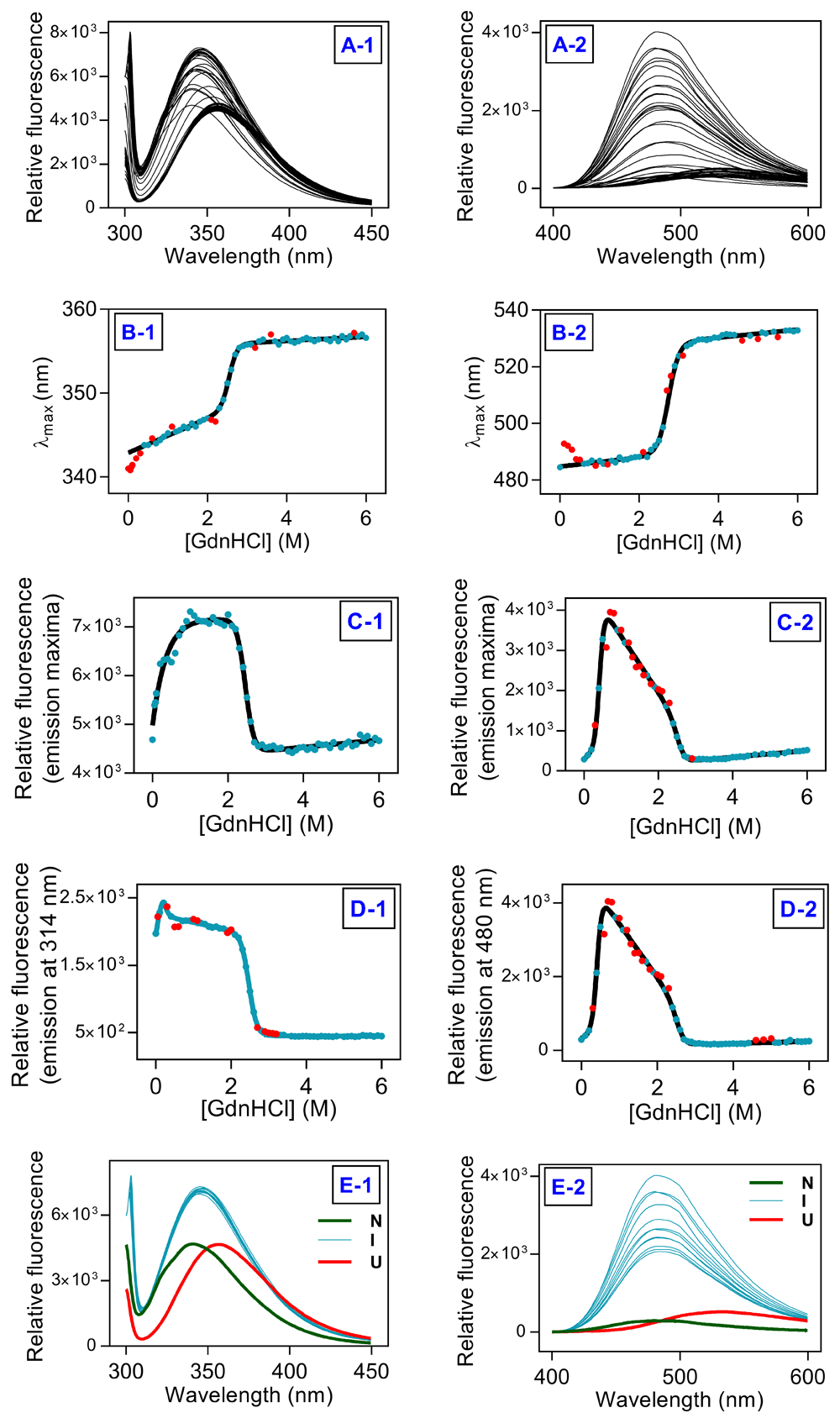
(A1) Trp fluorescence scans and (A2) ANS fluorescence scans. (B1) λmax (Trp fluorescence) and (B2) λmax (ANS fluorescence) against [GuHCl]. (C1) Imax (Trp fluorescence) and (C2) Imax (ANS fluorescence) against [GuHCl]. (D1) I295/314 (Trp fluorescence) and (D2) I370/480 (ANS fluorescence) against [GuHCl]. (E1) Trp fluorescence and (E2) ANS fluorescence of samples in native (green), intermediate (cyan) and unfolded state (red). Solid lines represent fit to the unfolding transitions, filled symbols represent data points from unfolding experiments, red symbols represent outliers.
Unfolding profiles of hAR equilibrated at different denaturants concentrations in far-UV CD are presented in Figure 4A1 and 4A2 for GuHCl and urea, respectively. Thermodynamic stability of hAR was determined on the basis of three state model by plotting change in ellipticity at 219/222 nm as a function of denaturant concentration (Figure 4B1 and 4B2 for GuHCl and urea, respectively). The transition determined by far-UV CD detected intermediate state at similar concentrations of denaturant as interrogated by fluorescence spectroscopy. All three states can be clearly distinguished from Far-UV CD profiles (Figure 4C1 and 4C2 for GuHCl and urea, respectively). Thermodynamic parameters derived from far-UV CD data are listed in Table 1.
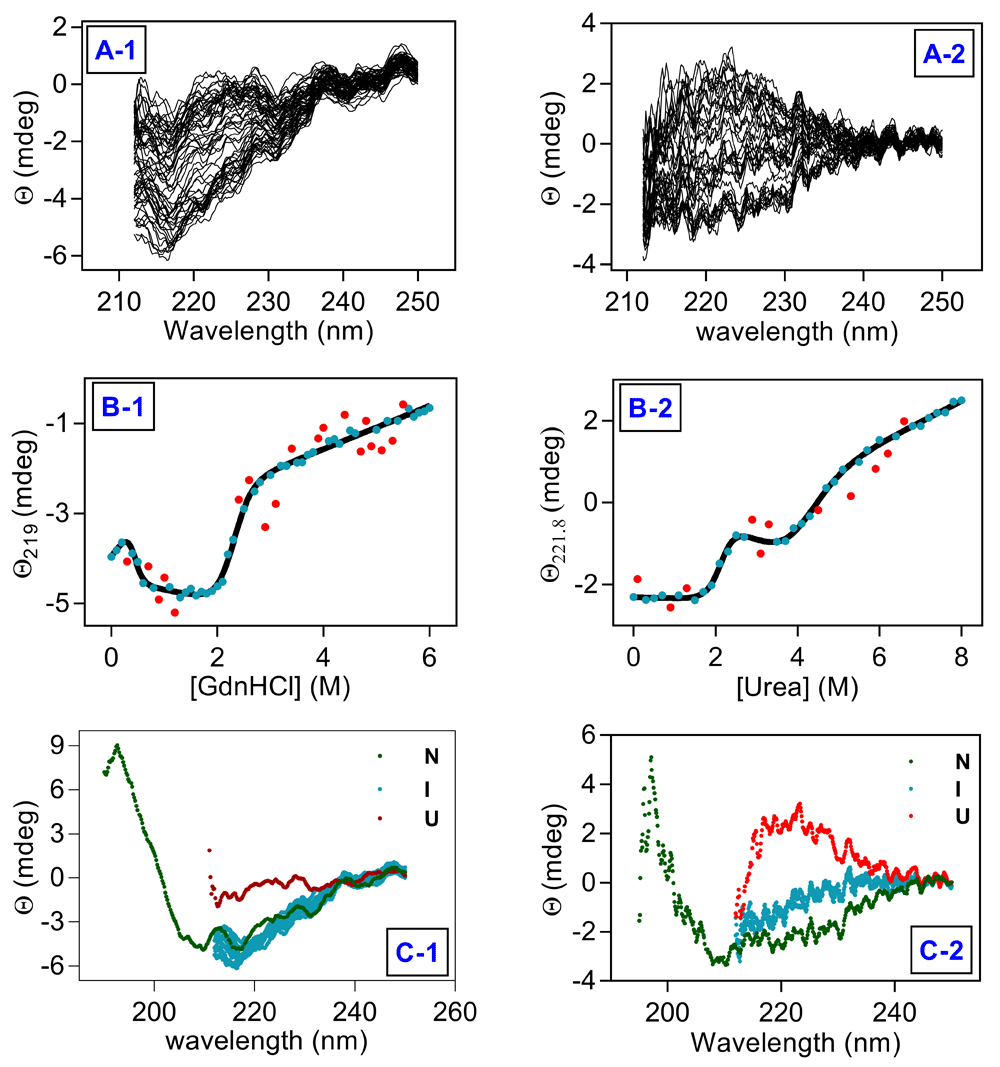
(A1) far-UV CD scans recorded for GuHCl and (A2) far-UV CD scans recorded for urea for all the samples. (B1) change in ellipticity at 219 nm against [GuHCl] and (B2) change in ellipticity at 221.8 nm against [urea]. (C) CD spectra of samples in native (green), intermediate (cyan) and unfolded state (red) for (C1) GuHCl and (C2) urea, respectively. Solid line represents fit to unfolding transitions, filled symbols represent data points from unfolding experiments and red symbols represent outliers in data fitting.
The intermediate state with enhanced ANS fluorescence and significant blue shift of λmax pointed to an intermediate state with some sort of ‘molten’ nature during hAR unfolding. Far-UV CD studies strongly suggest that the intermediate state retains significant secondary structure during urea- and GuHCl-induced unfolding.
During chemical induced unfolding hAR unfolds through an intermediate state which is absent during thermal unfolding. Moderate concentration of denaturant is known to stabilize native or intermediate state17. Absence of such stabilizing agent may be the reason that the intermediate state was not detected during thermal unfolding.
In all three probes used in studying unfolding, value of ΔG(N-I) obtained is ~30 kJ mol-1 and ~15 kJ mol-1for urea- and GuHCl-induced unfolding respectively while a ∆Gs of ~70 kJ mol-1 is almost same for both denaturants (Table 1). Thus, while urea seems to stabilize the native state with respect to the intermediate state, GuHCl seems to stabilize the intermediate state with respect to the native state.
It is known that small molecules change the free energy landscape of protein upon binding by selectively stabilizing native or intermediate/unfolded state18. Difference in value of ∆Gs obtained from thermal and chemical induced unfolding is ~20 kJ mol-1 (Table 1), which is most likely due to free energy of stabilization and destabilization by urea and GuHCl, respectively.
Values of ΔG(N-I) obtained from analysis of ANS fluorescence data are 16.48 and 28.61 kJ mol-1 for GuHCl and urea, respectively (Table 1), which indicate that intermediate state is not separated by a steep energy barrier from native state. Values for ΔG(I-U) obtained from ANS fluorescence are 57.26 and 38.6 kJ mol-1 for GuHCl and urea respectively (Table 1), which indicate that the intermediate state is separated from unfolded state by a high energy barrier. Thus, the intermediate state of hAR is close to its native state which makes it functionally more relevant.
In summary, equilibrium unfolding studies of hAR have led us to discover that hAR unfolds through an intermediate state, which is close to native state, and might have physiological relevance under hyperglycemic conditions in diabetes.
Figshare: data_f1000_hAR_unfolding.zip. https://doi.org/10.6084/m9.figshare.8001998.v115.
This project contains raw data for chemically and thermally induced unfolding studies on human aldose reductase.
Data are available under the terms of the Creative Commons Attribution 4.0 International license (CC-BY 4.0).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)