Keywords
BSEP/ABCB11, ABCB11-KO, Insilico, upregulation, HepG2, glibenclamide, ATP, and metformin, Nuclear Receptors
BSEP/ABCB11, ABCB11-KO, Insilico, upregulation, HepG2, glibenclamide, ATP, and metformin, Nuclear Receptors
The bile salt export pump (BSEP), the major bile salt transporter in the liver canalicular membrane, is coded by ABCB11 gene, and mutations in this gene cause progressive familial intrahepatic cholestasis type- 2 (PFIC-2)1,2. Besides PFIC-2, mutations or insufficiency of BSEP is associated with a variety of diseases such as drug-induced cholestasis, pregnancy induced cholestasis, cryptogenic cholestasis, cholangiocarcinoma and hepatocellular carcinoma, which are cancers of the liver3–7. Naturally, ABCB11 expression is induced by bile salts and is mediated by FXR- RXR heterodimer8. Here in this pilot study we explored in silico the interactions/networks around ABCB11. We wanted to identify the genes, drugs, microRNAs which might influence the expression of ABCB11. Drugs which could upregulate ABCB11 expression may be useful in ABCB11 haploinsufficiency and inhibition of the pump could result in the accumulation of toxic bile salts inside hepatocytes. Modulation of ABCB11 expression could be clinically beneficial in a variety of medical conditions.
We analyzed the microarray data set of the liver samples from ABCB11 knockout mice (GEO accession GSE70179) using GEO2R online tool from NCBI9. All differentially expressed genes (DEGs) were filtered with two criteria: -1> log2FC >+1 and adj. p-value <0.05.
To identify DEGs which are significant, pathway enrichment analysis was conducted using the ClueGo v2.5.5 app from Cytoscape10. ClueGo constructed and compared networks of functionally related GO terms with kappa statistics, which was adjusted at >0.4 in this study.
The protein-protein interaction (PPI) networks were built by the Search Tool for the Retrieval of Interacting Genes (STRING v11.0)11 and Cytoscape v3.7.1 software. The Molecular Complex Detection (MCODE v1.6), app from Cytoscape was used to screen modules of the PPI network with degree cut-off = 2, node score cut-off = 0.2, k-core = 2, and maximum depth = 100. The hub genes were identified by the CytoHubba v0.1 app. The top 10 nodes were considered as notable hub genes and displayed.
CyTargetLinker v4.1.0 from Cytoscape was used to identify the transcription factors (TFs) and microRNAs using ENCODE and Target-scan databases, respectively. We drew Homo sapiens TF-target interactions linkset from database (ENCODE)12 and drug-target interactions linkset from the database (DrugBank)13. The networks were visualized and analyzed using Cytoscape v3.7.1 Cytoscape app CyTargetlinker version 4.1.0[6] was used to screen the transcription factors and microRNAs
HepG2 cells were grown in high-glucose DMEM (Hi-Media Lab, Mumbai, Cat. # AL111-500ML) supplemented with 10% fetal bovine serum (CellClone, Genetix Biotech Asia, New Delhi, Cat.# CCS-500-SA-U), penicillin and streptomycin (Hi-Media, Mumbai Cat. # A018-5X100ML). When cells became 80% confluent, they were individually treated with glibenclamide (500 ng/mL)14, metformin (25 mg/L)15 or ATP (1 mM) for 48 h. After 48 h cells were scraped out for total protein and RNA.
Total proteins from HepG2 cells were prepared and run on 10% SDS-PAGE and transferred to a PVDF membrane using a transfer apparatus following the standard protocols (Bio-Rad). After incubation with 5% nonfat milk in TBST (10 mM Tris, pH 8.0, 150 mM NaCl, 0.5% Tween 20) for 1 h the membrane was washed once with TBST and incubated overnight at 4°C with rabbit antibodies against human ABCB11 (Affinity, Catalog #DF 9278) 1: 2000 dilution; mouse anti-human β-actin (Santa Cruz Cat.# SC4778), dilution 1:1000. The membrane was washed three times (TBST) and incubated with a 1:5000 dilution of horseradish peroxidase-conjugated anti-rabbit (Santa Cruz Cat# SC-2004)/anti-mouse antibodies (Cat.#SC-2005) for 2 h. Blots were washed with TBST four times and developed with the ECL system (Bio-Rad, US Cat.#170-5060) according to the manufacturer's protocol. The western blot images were acquired using iBright CL1000 (Invitrogen, Thermo Fisher Scientific).
Total RNA was isolated using NucleoZOL (Takara Cat. No. 740404.200) following manufacturer's instruction. cDNA was prepared from (deoxyribonuclease treated) total RNA using RevertAid Reverse Transcriptase (Thermo Cat. No. EP0441) following the manufacturer's instructions. Real Time PCR was done with unique oligonucleotide primers targeting ABCB11 and GAPDH, Ta=60°C, in triplicates and two repeats, using GoTaq® qPCR Master Mix (Promega Cat. No. A6001) following 'manufacturer's instructions on a Veriti Thermo Cycler from Applied Biosystems Waltham, Massachusetts, USA and data was acquired using the software associated with the same machine (ViiA7 V1.2) and relative quantification was calculated using the by 2(–ΔΔCt) method. Oligonucleotide primer sequences are listed in Table 3.
An earlier version of this article can be found on biorxiv.org (DOI: https://doi.org/10.1101/2020.09.01.277434).
Gene expression profile ABCB11 knockdown dataset GSE 70179 from GEO datasets were analysed with GEO2R tool. Genes with >2-fold change in expression value and <0.05 adjusted p-value was filtered. Identified differentially expressed genes (DEG) from the GSE dataset were classified in two groups - upregulated (375 genes) and downregulated (185 genes) (Extended data, Supp.Table-1)36. Gene ontology analysis was performed for functional analysis of DEGs by using ClueGo app from Cytoscape. PPIs of DEGs were constructed using STRING database showed an upregulation of genes related to cellular transport (pink colored nodes), and these nodes were also shared by Toll-like receptor (TLR) signalling (Figure 1). Downregulated genes were involved in metabolic pathways (sterol, carbohydrate, alcohol, etc.) (Extended data, Supp. Table-2)36. We next identified top hub genes in PPI network using CytoHubba app from Cytoscape (Table 1). Immunologically important genes were among the top ranked upregulated hub genes (Figure 2a) downregulated group majorly represents cell signaling and fatty acid metabolism (Figure 2b). Epidermal growth factor receptor (EGFR) ranked first among the genes involved in signaling pathways. Kinases play a role in the transcription, activity, or intracellular localization of ABC transporters as do protein interactions16. Proteins interacting with ABCB11 are represented in Figure 3 which includes nuclear receptors NR1H4 and NR0B2. Most proteins were associated with bile acid metabolism and transport.
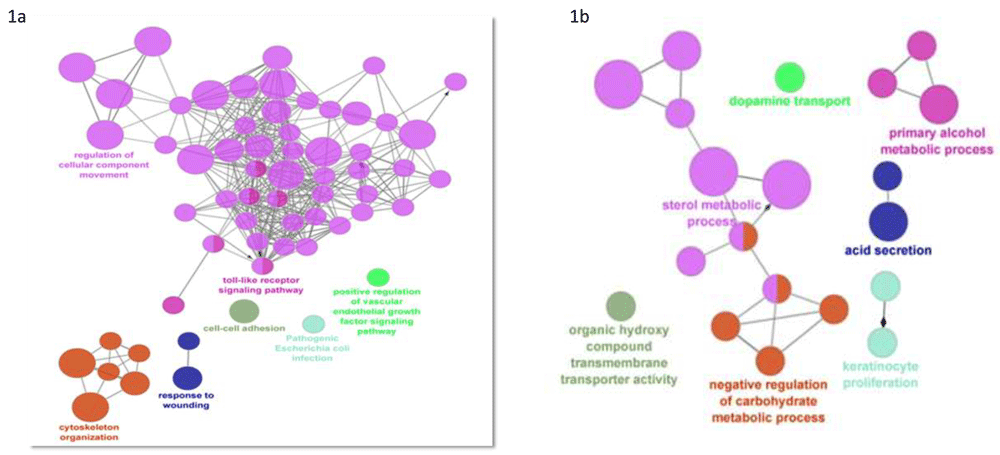
Gene ontology analysis was performed for functional analysis of DEGs by using ClueGo app from Cytoscape. This app allows simultaneous analysis of multiple annotation and ontology sources. Functionally grouped network is represented Figure 1a (upregulated genes) 1b (downregulated genes) . The node size represents enrichment significance and connections are based on kappa score (> 0.4). In upregulated gene group maximum number of nodes which are in pink color represent the cellular component movement. These nodes are shared by toll like receptor signaling pathway.
We observed that the top ranked hub genes in PPI network which were upregulated were associated with immune activity while those downregulated are associated with cell signaling and fatty acid metabolism. EGFR came first in the ranking which is a critical receptor in several cell signaling pathways.
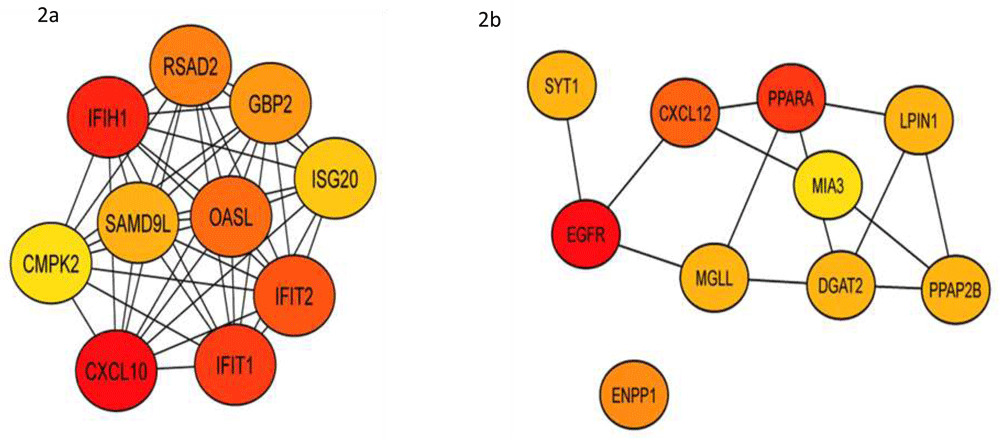
We observed that the top ranked hub genes in the upregulated group were mainly related to immune activity (a). The top hub genes in downregulated group were associated with cell signaling and fatty acid metabolism. EGFR emerged as the top hub gene, a growth factor receptor which is crucial factor several cell signaling pathways (b).
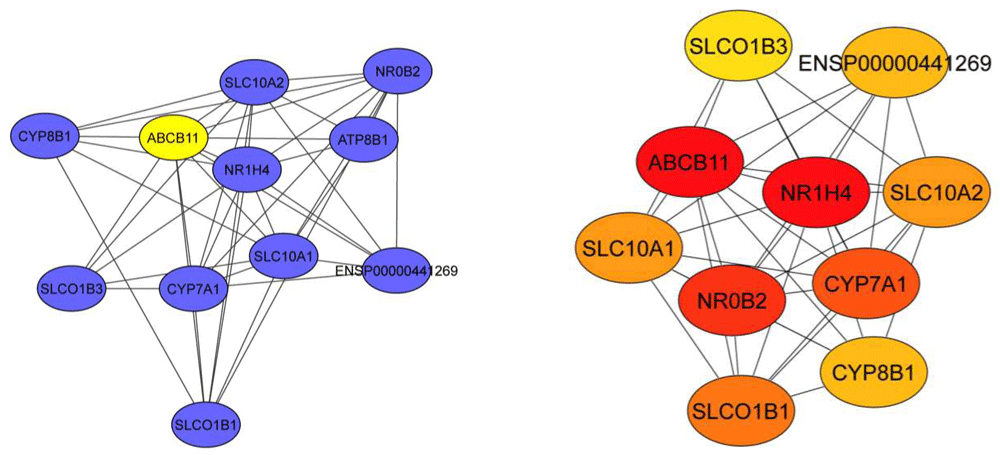
This network was constructed to analyze the relationship between ABCB11 and other proteins. Cytohubba app was used to calculate centrality of each node by MCC method. Node colour (red to yellow) represents the significance of the centrality in the group. In this analysis, we counted 11 nodes and 42 edges. These proteins majorly involved in bile acid metabolism and transport. Most of these genes are participant of more than one pathway which was expected because these pathways intersect and coregulated. We also mapped the NR0B2 protein, which is participate in sterol metabolism.
As described, sub-network analysis was performed using MCODE (Figure 4), and CMPK2, ACTG1, and SSTR2 emerged as seed nodes among upregulated genes (Table 2). Among downregulated gene groups, only one subnetwork was found to be significant which had three genes: MIA3 (which codes a protein which is important in the transport of cargos that are too large to fit into COPII-coated vesicles such as collagen VII), IGFBP4 (encoding a protein that binds to both insulin-like growth factors and modifies their functions) and NOTUM (encoding a carboxylesterase that acts as a key negative regulator of the Wnt signaling pathway by specifically mediating depalmitoleoylation of WNT proteins).
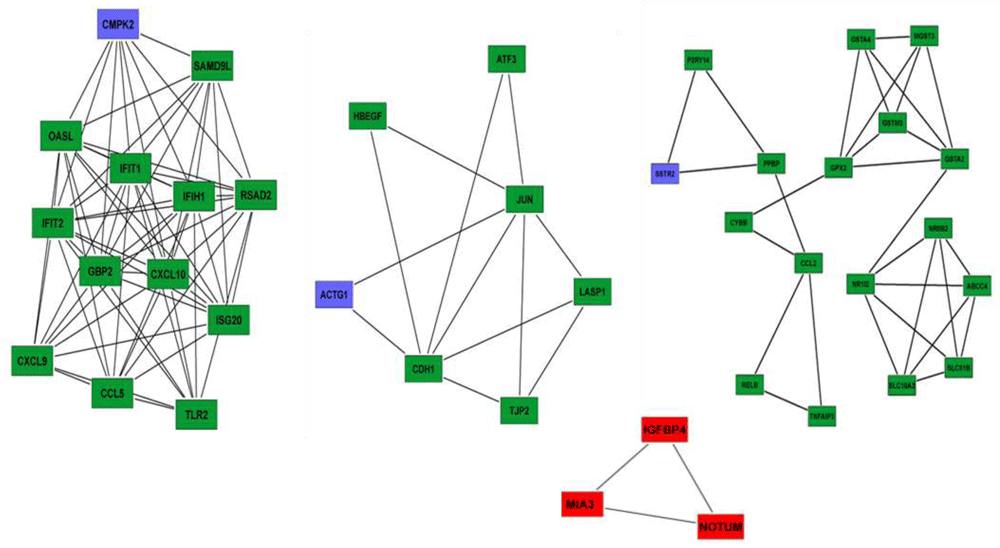
Top sub-networks on the basis of MCODE score (Degree cut-off= 2, node score cut-off = 0.2, k-core = 2 and max. depth = 100). Upregulated gene group clusters, we identified seed nodes (CMPK2, ACTG1 and SSTR2) in the network (green and blue). In downregulated gene group, we identified only one subnetwork which qualified cut off criteria. Three genes in this sub-network was identified: MIA3, IGFBP4 and NOTUM (red).
Sub-network analysis was performed using the Molecular Complex Detection (MCODE) app from Cytoscape and CMPK2, ACTG1 and SSTR2 emerged as seed nodes among upregulated.
| Gene | Forward | Reverse |
|---|---|---|
| GAPDH | GAAGGTGAAGGTCGGAGT | GAAGATGGTGATGGGATTTC |
| ABCB11 | CCTCCATCCGGCAACGCT | CACTGAATTTCAGAATCCTCCTAACTGGG |
Using CyTargetLinker identified two drugs, glibenclamide, and ATP, directly targeting ABCB11. We subsequently looked for microRNAs [Target-scan database]17 that were associated with ABCB11, and a total of 14 microRNAs were identified targeting the gene (Figure 5). Transcription factors and microRNAs targeting ABCB11 and interacting partners are represented in Figure 6.
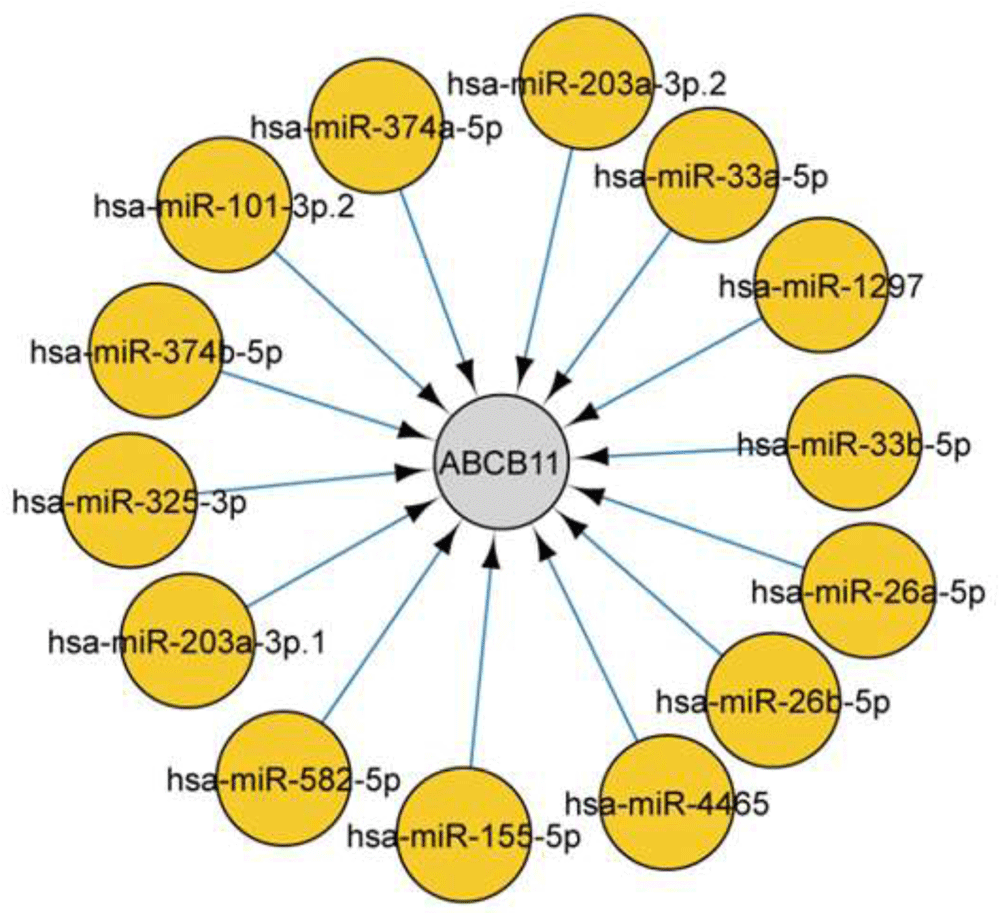
We identified microRNAs that were associated with ABCB11. In total 14 microRNA identified targeting the gene.
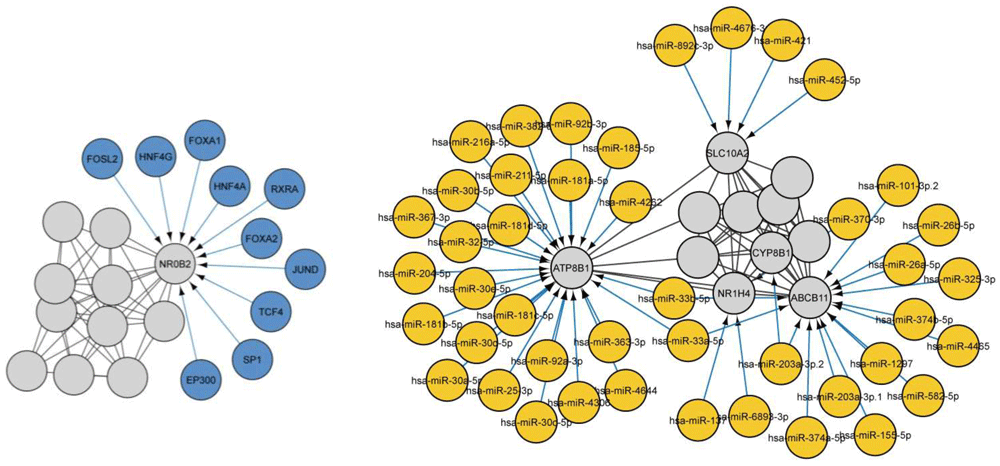
In the screen of transcription factors of ABCB11 interaction network we observed 21 nodes and 52 edges. Among these transcription factors, FOXA have been suggested an important factor in bile duct development and lipid accumulation. HNF4A in the regulation dyslipidaemia and terminal liver failure and JUND in fibrosis development. Others can be investigated in future studies. We counted 55 nodes and 89 edges in the search of microRNA targeting the ABCB11 network. Four genes (ABCB11, ATP8B1, SLC10A2 and NR1H4) targeted by multiple microRNAs also some microRNA such as has-miR-203a-3p.2 and has-miR-203a-3p.2 target more than one gene. By nature, a microRNA can regulate several pathways therefore it would be interesting to study in future the dysregulation of these microRNAs and interaction with Identified transcription factors.
We evaluated in vitro, the effect of three drugs, two of which were bioinformatically predicted (Glibenclamide, ATP) and one based on literature18. We found all the three compounds upregulating ABCB11 expression based on qPCR, and this was confirmed by western blot (Figure 7). Unannotated western blot images and raw qPCR Ct values are available as Underlying data36.
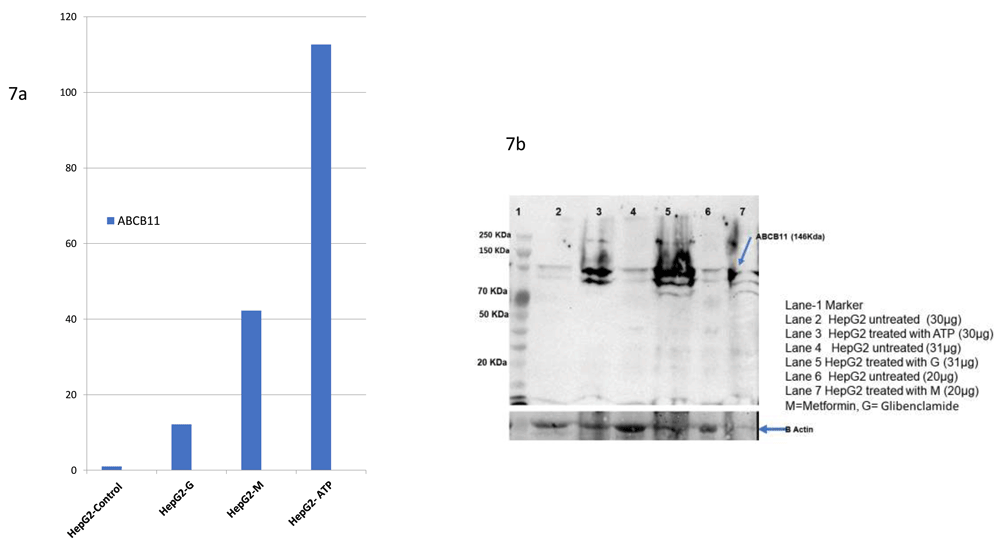
All the three compounds upregulated ABCB11 expression based on Real Time PCR data. This was further confirmed by Western Blot against anti-human ABCB11 antibody as the primary antibody. The upper image in 7b shows the PVDF membrane probed with antibody against ABCB11 and the lower image shows the same membrane stripped and probed with antibody against beta-actin (loading control).
We identified several immunologically important genes being upregulated during ABCB11 deficiency. The reason could be liver cell injury secondary to bile salt accumulation, which triggers the sterile immune response19,20 and the downregulation of transport proteins and metabolically important genes could be because of decreased liver function following damage. A regenerative response follows cell injury, and a host of genes involved in regeneration are upregulated21–23; however, it appears that bile salts in the absence of BSEP hamper the regenerative response reflected by dysregulated collagen transporting protein MIA3 and NOTUM a protein involved in Wnt signaling. It's also possible that EGFR is dysregulated via accumulating bile salts mediated by STAT324. We have observed an upregulation of ABCB11 in a liver cell line (HepG2) on treatment with glibenclamide, metformin, and ATP. This expression is upregulation may be a compensatory mechanism in the case of glibenclamide and metformin because these drugs are known to inhibit ABCB1125. Metformin is known to interfere with ABCB11 function, mediated through AMPK-FXR crosstalk18 involving metformin induced FXR phosphorylation. ATP acts through ATP receptors on hepatocytes26,27. ATP is known to cross the plasma membrane28 and this can act via AMPK. However, ATP has a very short half-life29, and it may be converted to ADP, which can activate AMPK30. In a recent report, metformin was shown to suppress ABCB11 expression, which is not in agreement with our observation, however, they performed their experiment on primary human hepatocytes, and they have also treated their cells with dimethylsulfoxide (DMSO)31.
There are many reports stating the influence of DMSO on human gene expression. For example, Verheijen et al. “exposed 3D cardiac and hepatic microtissues to medium with or without 0.1% DMSO and analyzed the transcriptome, proteome and DNA methylation profiles”. They found that “in both tissue types, transcriptome analysis detected >2000 differentially expressed genes affecting similar biological processes, thereby indicating consistent cross-organ actions of DMSO”. In both tissue types, the transcriptome analysis detected over 2000 differentially expressed genes affecting similar biological processes32. Moskot et al. reported alterations of lysosomal ultrastructure upon DMSO treatment33. Alizadeh et al. reported that DMSO catalyzes hepatic differentiation of adipose tissue-derived mesenchymal stem cells34. It has been observed that “culturing pluripotent stem cells in DMSO activates the retinoblastoma protein, increases the proportion of cells in the early G1 phase of the cell cycle, and subsequently improves their competency for directed differentiation into multiple lineages in more than 25 stem cell lines”35. However, we are not sure whether the observed difference is attributed to DMSO.
In conclusion, we need more experiments to determine the mechanisms of action of these drugs on the upregulation of ABCB11. Many changes in gene expression following ABCB11 knockout could be secondary to stress, immune and regenerative responses following hepatocyte injury in mice liver.
Harvard Dataverse: Real Time PCR for ABCB11 and few NRs. https://doi.org/10.7910/DVN/AOYKY736.
This project contains the following underlying data:
2020-09-12 092712-ViiA7-export.xls. (qPCR data following addition of ATP, metformin or gilbenclamide.)
ABCB11 WESTERN BLOT DRUG.tif. (Unannotated western blot image for ABC11.)
ABCB11_WB_Repeat_Drugs.tif. (Unannotated repeat western blot image for ABC11.)
actin drug.tif. (Unannotated western blot image for β-actin.)
Actin_2020_07_11_182456.jpg. (Unannotated western blot image, including β-actin loading control.)
Actin_2020_07_11_182456.tif. (As above, but in tif format.)
nisha_qPCR DATA_ 7142020.xls. (qPCR data for ABC11 and other indicated genes.)
realtime and western blottt (1).pptx. (Western blot and qPCR data pooled into a single file.)
Repeat_Actin_drug_WB.tif. (Unannotated repeat western blot image for β-actin.)
Harvard Dataverse: Real Time PCR for ABCB11 and few NRs. https://doi.org/10.7910/DVN/AOYKY736.
This project contains the following extended data:
Supp-Table-1-Dysregulated genes GSE70179, GEO2R, NCBI. (Differentially expressed genes
Supp-Table-2-Gene ontology analysis, DAVID. (Gene ontology analysis of
Data are available under the terms of the Creative Commons Zero "No rights reserved" data waiver (CC0 1.0 Public domain dedication).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)