Keywords
Combined workflow, ImageJ2, FIJI, cell shape modelling, R-statistics, machine learning, clustering, image processing, image analysis, automation, drug screening
This article is included in the Bioinformatics gateway.
This article is included in the NEUBIAS - the Bioimage Analysts Network gateway.
Combined workflow, ImageJ2, FIJI, cell shape modelling, R-statistics, machine learning, clustering, image processing, image analysis, automation, drug screening
Typos corrected. Questions regarding the workflow where updated in figure 3 to address the reviewer questions. Raw data was added to the osf repository. Figures are annotated according to the image type (brightfield or fluorescence micrograph) that was used. Figure 3 was updated.
See the authors' detailed response to the review by Janos Kriston-Vizi and Qing Hsuan Ong
See the authors' detailed response to the review by Massimiliano Lucidi
Bioimage analysis is continuously changing our understanding about the world and how we see our environment. Bacteria are present at µm scale and physiological processes, like cellular signalling events, are even lower than nanometer scale. Cell shape is important for these non-compartmentalized, unicellular organisms. The question of how fast cells grow and divide is connected to tightly regulated intracellular processes like protein-biogenesis from which novel synthesized proteins are translocated along synthesis pathways to their target. Associated proteins play an important role in membrane biogenesis1–3. Membrane proteins are synthesized by ribosomes and usually co-translationally inserted into the cytoplasmic membrane. In this process, the signal recognition particle (SRP; composed of SRP RNA and of Ffh protein) recognizes the signal sequence of the nascent polypeptide at the ribosome4. This complex is recognized by the SRP receptor FtsY and is delivered to the translocon in the cytoplasmic membrane5–7. The nascent polypeptide can be inserted into the membrane or translocated across the membrane by the translocon8,9. Disruption of these processes results in dysregulation of essential networks followed by reduced viability, for instance, via chemically induced stress by antibiotic compounds. Susceptibility towards different antibiotics can vary from organism to organism depending on the mode of action of the compound. To further understand respective mechanisms of cellular stress response in bacteria, morphological feature changes are useful to monitor those in vivo. Our goal is to illustrate how an imaging based workflow can be efficiently deployed to monitor these processes as part of an antibiotic drug screening strategy involving cell morphology and viability. Furthermore, the presented combined workflow shows how to extract valuable information from imaging data in a reproducible manner using classic statistical approaches, as well as unsupervised machine learning algorithms.
Our model organism Shewanella putrefaciens CN-32 is a Gram-negative bacterium that occurs in aquatic environments10. Depending on its growth and division cycles, it has approximately 3 µm in length and 1 µm in width at exponential phase (see Figure 1). Cell division occurs with peak rates at exponential phase represented by OD600 0.5. In our study, we use markerless insertions at the original gene locus functionally expressing fusion-proteins that are relevant for membrane protein-biogenesis: bacterial signal recognition particle Ffh, its receptor FtsY and ribosomal protein of large subunit L1. In order to monitor drug induced stress responses that affect protein-biogenesis, mVenus is used as a fluorescent protein for fluorescence microscopy. Recombinant strains are cultured at 30°C and 200 rpm in Lysogeny broth (LB) medium without antibiotics. This aspect is beneficial to avoid potential bias induced by compound interaction. To analyze antibiotic stress on membrane biogenesis, the protein synthesis inhibitor puromycin was used (200 µg/ml). Puromycin inhibits translation via early termination and subsequent premature release of the nascent polypeptide chain (reviewed by Aviner in 2020)11,12. Additionally, RNA-polymerase inhibitor rifampicin (RIF)13 (25 µg/ml) and peptidyl transferase inhibitor chloramphenicol (CM)14 (50 µg/ml) were used to confirm outcomes of the deployed workflow15.
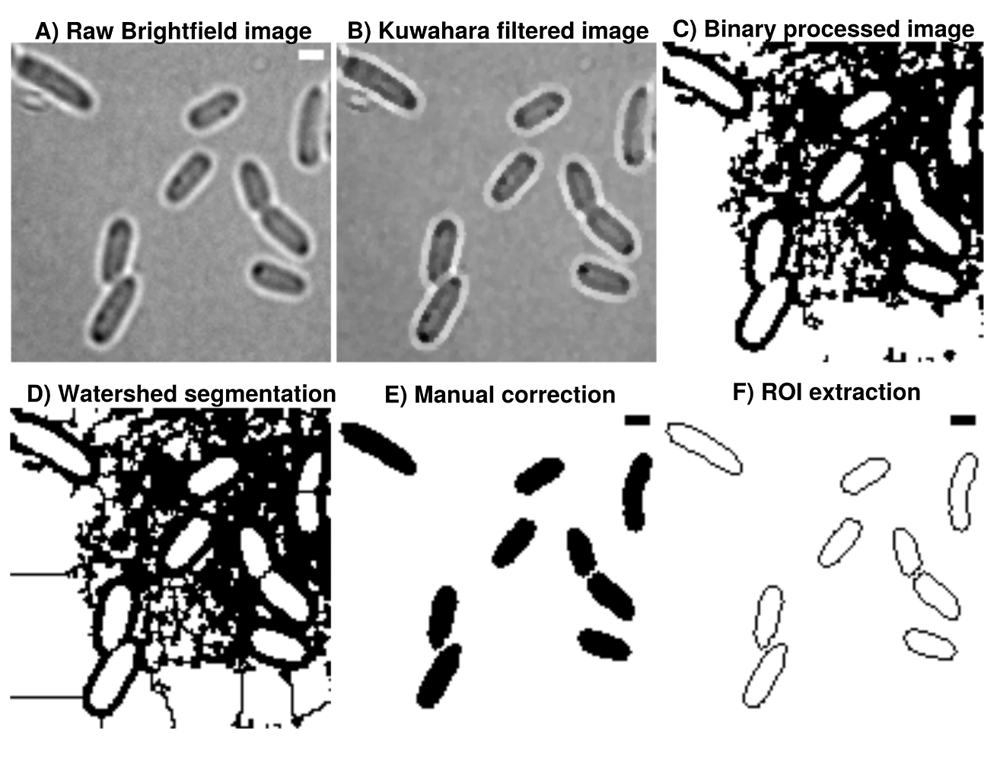
A) Brightfield images are transformed from 16-bit into 8-bit and smoothed by B) non-linear noise reduction using the Kuwahara filter option in FIJI (sampling window = 2). Noise reduced images are C) thresholded using ’Percentile’ thresholding algorithm and D) segmented using watershed for cell discrimination. E) Resulting binary images are finally corrected manually by correcting potential false positive cells through drawing options. F) ROI are extracted from appropriate binary images, stored as individual .zip folders which also involves measurement of cellular areas in an automated manner. This process can be repeated until the the correction is optimal and representative for further analysis. Further analysis always refer to the updated data base and corrections are automatically included when image analysis is reproduced accordingly in Celltool or R-statistics. Scale bar = 1 µm.
Drug screening time-course replicates are taken at different days. Cells are inoculated from an overnight culture and cultured at 30°C and 200 rpm until OD600 0.5. From this batch, 1 ml is sampled into a glass tube as steady-state (NC), 1 ml treated with antibiotics and is continuously incubated with same conditions. A sample of 3 µl is taken from the culture at steady state and mounted on slides using 1% ultrapure agarose for reduced background. During imaging of cells at steady-state, cells treated with antibiotics are incubated for a minimum of 30 minutes until image acquisition. Similar to this, samples from treated culture after a minimum of 60 minutes are taken accordingly. Viability assay is additionally performed after timecourse experiments. We assume that cells treated with the compound during the time-course experiment are sublethally impaired in cell-growth and division. In order to do so, cells at steady-state and treated with puromycin are 10 fold serially diluted and distributed on LB-agar plates for further incubation at 30°C overnight and imaged using a Fusion-Gel-Illuminator.
Images are acquired using brightfield and fluorescence microscopy. In our live-cell imaging pipeline, we use an Olympus IX 71 microscope (100x/NA 1.49/optovar 1.6) customized for slimfield microscopy16 with a fast image acquisition conducted by an Andor iXON Ultra EMCCD camera. Brightfield images are acquired using 50 ms exposure time (see Figure 1). Live-cell time-lapse recordings are acquired using 16 ms exposure time. We take advantage of photobleaching steps through continuous slimfield illumination until single particles can be localized (see Figure 2). Continuous slimfield excitation for photobleaching of the samples is conducted using 514 nm laser line (50%). The microscope setup uses Andor Solis as camera software using at least 2000 frames with integration times of 17.76 ms. Further information about the raw data used in this study is covered in Mayer et al., 202115.

In contrast to more advanced imaging techniques like phase contrast and Nomarski-microscopy, regular brightfield images are more difficult to process into binary images and resulting ROI possibly need manual correction (green arrow). The A.D.I.C.T. workflow is capable of processing binary images that can be segmented further using watershed segmentation as shown with this study. Although images need to be manually corrected due to the limitation of the imaging technique, cells are thresholded in a more conservative manner due to the use of pixel preserving thresholding algorithm 'Percentile'. 'Otsu' introduces artificial gaps more frequently inside ROI (compare cyan and magenta arrows). Therefore, ‘Percentile’ is used for this data because it requires less correction. However, 'Otsu' thresholding algorithm is used in the paper to mask fluorescent micrograph projections for measuring intensity.
Image processing in this workflow is conducted using ImageJ2/FIJI17–21. Raw grayscale images are sequentially and automatically annotated, transformed from 16-bit into 8-bit, non-linearily noise reduced using a Kuwahara filter and thresholded using ’Percentile’22 thresholding algorithm and segmented using watershed segmentation for cell detection23,24 (see Figure 1). ‘Percentile’ is used in this workflow thus initial analysis of processed binary brightfield images showed that this thresholding algorithm is more efficient compared to more common algorithms like ‘Otsu’ (see Figure 2). Resulting binary images are finally corrected manually using the paint function of ImageJ2/FIJI accordingly (see Figure 1). This process can be repeated until the correction is optimal and representative for further analyses (see Figure 3). Fluorescence time lapse recordings are automatically annotated, cropped after sufficient photobleaching steps and projected using the ’Standard deviation’ method. This method is used for tomographic representations and highlights areas of high fluorescent densities within a region of interest (ROI). Resulting fluorescence micrographs are background corrected using the math function ’subtract’. All images are automatically scaled and stored in a database management system that is connected through a set of predefined folder operations within the automation script. Regions of interest (ROI) are automatically detected using ImageJ2/FIJI and further processed using scripts based on macro language implemented in ImageJ2/FIJI (see Figure 1). ROIs are extracted from binary images using the ROI-manager plugin and used to create a mask for cell-measures of projected fluorescence micrographs based on the use of ‘Otsu’ thresholding25 (see Figure 3).
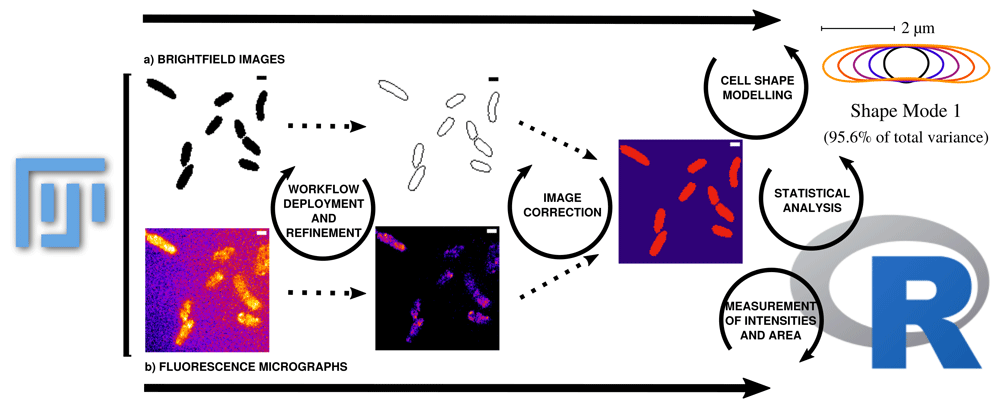
Data can be corrected and updated at every stage of the workflow to continuously improve the database until bias is minimized. a) ROIs are collected from binary images that are transformed from raw grayscale brightfield images. The measured area (µm²) using ImageJ refers to respective ROI and can be compared with the area results from Celltool (Celltool also calculates the area based on the transformed binary image). The measurements collected using the ImageJ function can be further used for area comparison in combination with mean gray values and integrated densities or can be applied as a mask to measure the fluorescence micrographs within the respective cell boundaries. This step eliminates signal that is located outside the ROI. b) Processing of the fluorescence time lapse recordings is conducted on the basis of cropping the relevant frames after sufficient photobleaching until single particle level is reached. Analysis of fluorescence micrographs is based on the measurements defined in ImageJ macros and then further processed using an R pipeline using the mentioned packages. Fluorescence micrographs that are used to measure intensities refer to the respective protein and cell vitality monitored over time. It is important to know that the raw data does not contain useful meta-data by scratch. The workflow enables the experimenter to fully control the annotation of the data in a reproducible way. The outcome should be that after the workflow (re-)deployment of the processing using the fluorescence micrographs, properly annotated data tables that can be combined for further analysis using R-packages are obtained. Intensity measurements directly correspond to protein levels and dynamics. Data can be therefore used to obtain more detailed perspectives about the observed biological process. Cellular stress can be monitored with different variables containing information for fluorescence activity and morphological changes during stress induction. To monitor the same data from different perspectives makes the analysis more robust and allows to explore the cellular stress response in a more detailed, simultaneous and semi-automated manner. This also helps to re-evaluate the analysis in a well documented manner thus the extraction of information and the processing starts from the raw data (brightfield and fluorescence micrographs). Further modeling for instance becomes easier thus data is well prepared for more complex analysis tasks. Taken together, the workflow allows to evaluate the observed cellular stress response at protein expression levels (fluorescence microscopy) in a simultanous and interconnectd (ROIs based on brightfield images) manner with stress response observed by cell shape abnormalities (Celltool). Scale bar = 1 µm.
As a result, we receive comma separated value (.csv) tables that are merged using a custom script in R-statistics based on the ’dplyr’ package to organize and merge the tables to a final result-table. Cell shape analysis is conducted using Celltool developed by Pincus lab for cell shape modelling26. Scripts for extraction of polygonal contours, alignment, principal component analysis (PCA), statistical evaluation of probability density of cell areas or curvatures and modelling of shape modes are adapted and modified according to the tutorial from Pincus labs (https://zplab.wustl.edu/celltool/). Statistical evaluation of cell areas collected from ROI and fluorescence measurements mean gray value (mgv) as well as integrated density (IntDen) (https://imagej.nih.gov/ij/docs/menus/analyze.html) are statistically analyzed with Rstudio (v.1.1.463) using a customized markdown pipeline in R 3.6.127–38 (see Figure 3). To understand the context between mgv and IntDen, it is important to know that:
Statistical distributions resulting from measurements are tested for normality using the Shapiro-Wilk test39. Non-parametric Wilcoxon rank sum test is used to test pairwise for significance (confidence level: 0.95 ; p < 0.05 = * ; p < 0.01 = ** ; p < 0.001 = ***)40,41. In order to establish an unsupervised machine learning approach using the ImageJ2/FIJI results tables, Density based clustering of applications with noise (dbscan) R-package is applied to cellular areas and mean gray value with the aim to evaluate clusters that distinguish between cellular amount of fusion proteins (indicated by fluorescence) and cellular areas42.
Puromycin treated cells show abnormal cell morphology regarding their size and shape over time (see Figure 4A). Cell shape changes can be modelled with Celltool. Generalized models according to time points show increased variation of cells explained by shape mode 1, 2 and 3 (see Figure 5), if stressed with puromycin. Corresponding to these findings, probability plots show increasing cell size (see Figure 4C) and abnormal cell morphology reflected by normalized curvature in a quantitative manner during induction with puromycin (see Figure 4D). During progression of the time-course, cell length increases which indicates cell division stress which was also shown by other groups before43,44. Corresponding to that, Celltool analysis under rifampicin and chloramphenicol induced stress shows similar outcomes (see Figure 6). Differences between cellular areas grouped by condition times show clearly a significant time dependent increase (see Figures 7A and B). Although differences of cellular areas appear to be significant as well between L1, Ffh and FtsY if grouped by respective strains, there is no significant difference between Ffh and FtsY. Furthermore, these findings are strongly influenced by extreme values suggesting that these differences are not a result due to puromycin (see Figures 7C and D). The fluorescence intensity of Ffh and FtsY decreases over time because no new proteins can be translated caused by the protein synthesis inhibitor puromycin and the old proteins are degraded (see Figure 7E and F). From here, it can be only speculated why L1 amount is higher after 30 minutes of induction. One possible explanation could be that the compound does not affect ribosome formation itself. Furthermore, the autocatalytic nature of the ribosomes makes it even more complicated to address45. However, the increased integrated density (see Figure 7F) indicates that changes of the cellular area (in 2D space of course, which is actually a volumetric aspect in 3D) is the decisive parameter to monitor puromycin stress response in our approach. Our assumption that the cellular area is indeed the relevant indicator for puromycin induced stress is further supported by the fact that comparison of cellular areas and curvature with Celltool corresponds to the R-statistics analysis.
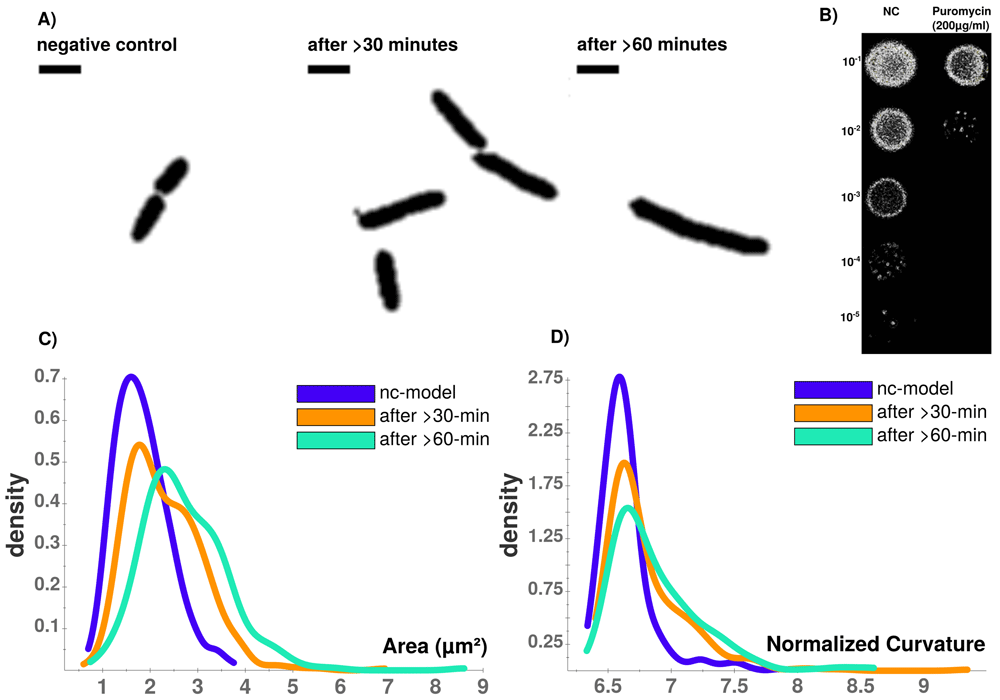
A) Time-course of puromycin stressed cells at different time points (NC, after >30 min., after >60 min. Viability assay with unstressed (NC) and puromycin stressed cells. B) Colonies on LB-agar show drastic effects between serial dilutions of steady-state and puromycin treated cells. Whilst viability is not impaired for steady-state, protein biosynthesis inhibitor puromycin shows sublethal impaired cell growth and division indicated by reduced colony density. C) Celltool area comparison shows increasing numbers of larger cells that can be quantified which corresponds to D) an increasingly abnormal cell curvature over time after puromycin induced stress. Scale bar = 2 µm.
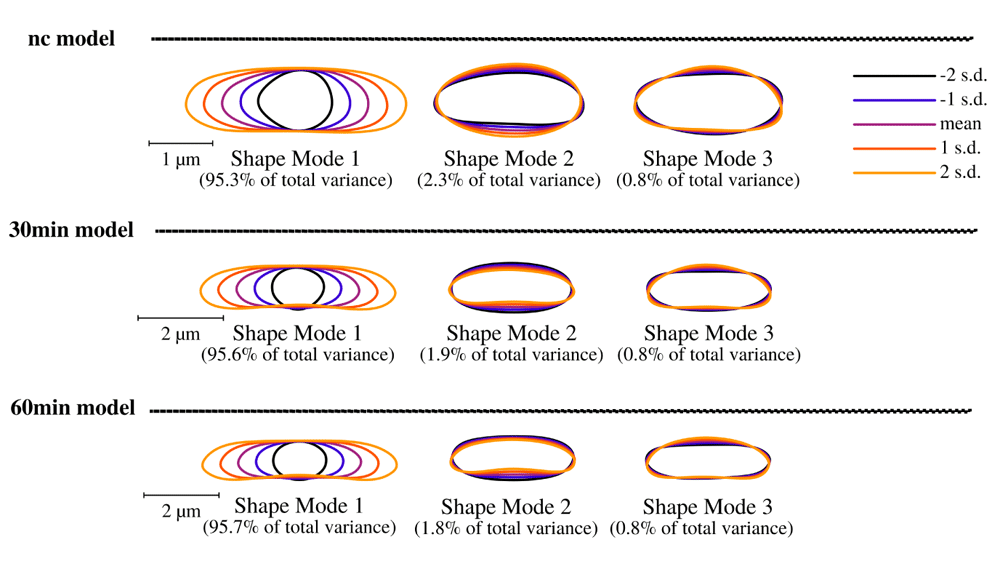
Brightfield image analysis using Celltool. Lower limits of explained differences of shape models caused by variation are taken into consideration if not lower than 0.8%. Our results suggest for all time-course acquisitions that mode 1 explains the majority of variation followed by width of the cells represented by mode 2 and 3. Considering the scale bar, the generalized models clearly show that puromycin is indeed affecting the cell length specifically thus cell division might be impaired. These findings correspond to the analysis of cellular areas and curvatures.
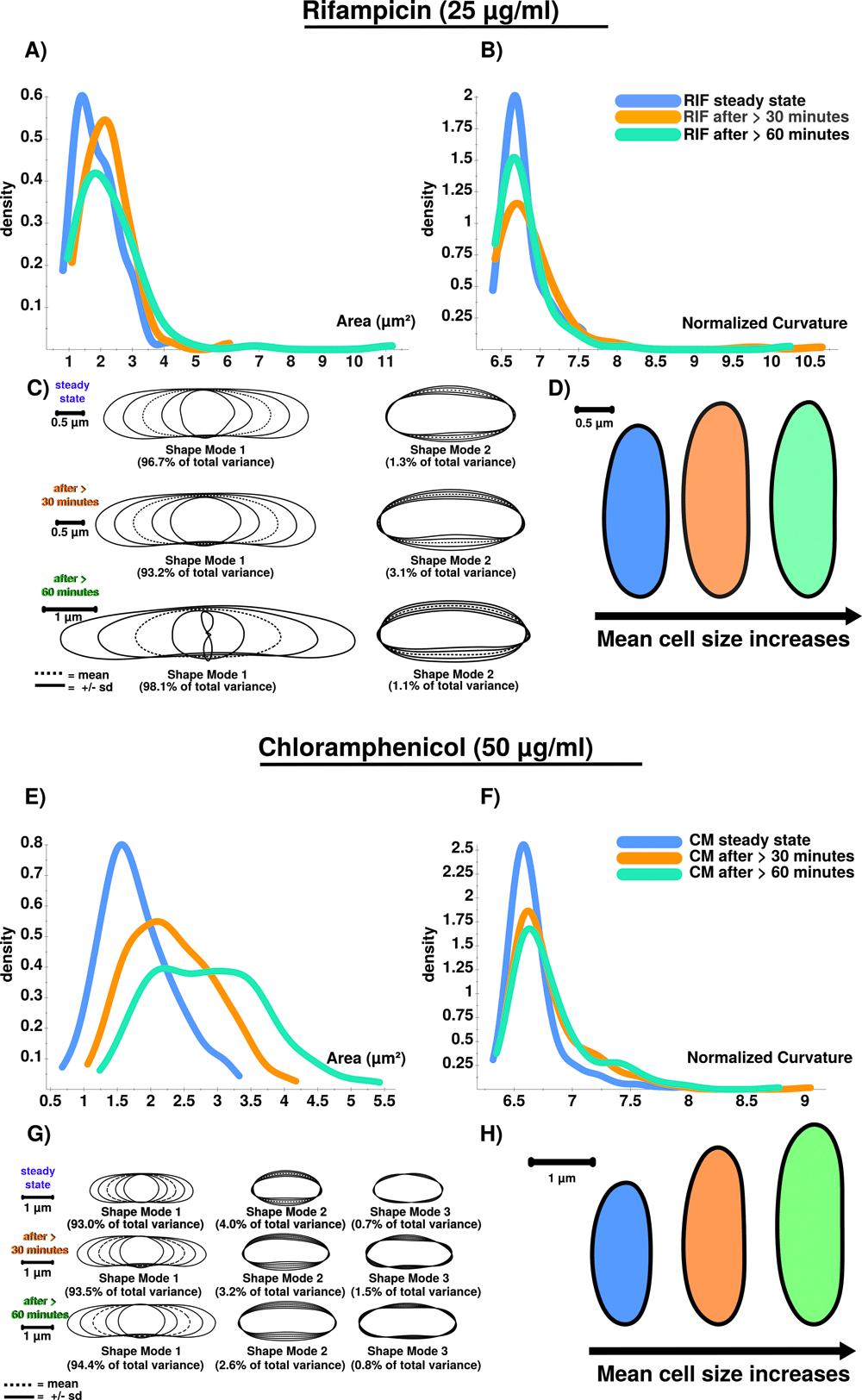
A) RIF stressed cells show increased cellular area over time. B) Curvatures for analyzed cellular populations are also increased after RIF stress is induced. However, curvature does only increase in total after stress induction, and not successively. Curvature is stronger affected after 30 minutes (orange) compared to conditions after 60 minutes (green), which could be triggered by an initial shock induced by RIF. C) PCA results confirm the stress response using shape mode 1 and 2. RIF stressed cells show a high variability regarding cell length. This is why the shape mode 1 after 60 minutes (green) is difficult to model (see the inner shape of mode 1 (-2 s.d.) ). Remarkably, RIF stress induces division stress according to our results, indicated by abnormally enlarged cells. D) Extracted and compared mean cell shapes indicate that RIF stressed cells tend to be larger compared to steady-state cells and show that the stress-response is detectable at 25 µg/ml. E) CM stressed cells show increased cellular area over time. F) Curvatures for analyzed cellular populations are increased after CM stress induction. Similar to RIF (see Figure 6B) curvature does only increase in total but not successively. G) PCA results confirm the stress response using shape mode 1, 2 and 3. CM induced stress results in a higher number of abnormally shaped cells. H) Extracted and compared mean cell shapes (mode 1) show that cells are successively enlarged during time courses compared to steady-state cells on average.
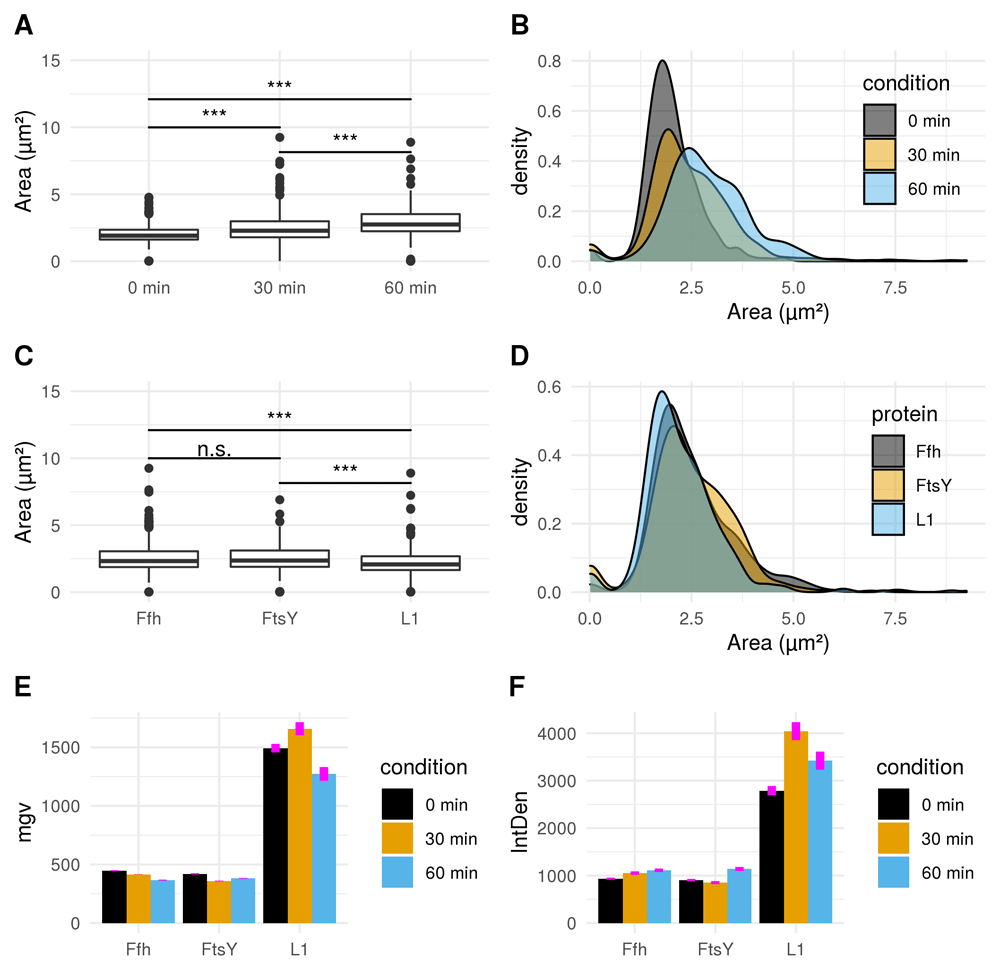
A) Boxplot comparison shows increased cell size distribution for all observed fusion proteins over time if stressed with puromycin (0 min: n = 412; 30 min: n = 455; 60 min: n = 338). Extreme value count increases over time for the pooled fusion protein samples. B) Probability density of larger cells increases over time for pooled fusion proteins. Results show decreasing number of cells with smaller area indicating that puromycin induced stress results in increase of larger cells for the whole sample. C) Cellular areas grouped according to respective strains containing different fusion proteins are differing: Ffh: n = 453; FtsY: n = 371; L1: n = 381. D) Probability density plot does not indicate a strong difference if compared between strains of different fusion proteins. E) Comparison of mean gray values (mgv) between groups of fusion proteins show slightly decreasing effects over time for Ffh (0 min: n = 131; 30 min: n = 184; 60 min: n = 138) and FtsY (0 min: n = 112; 30 min: n = 160; 60 min: n = 99). L1 (n = 169) appears to increase after 30 min (n = 111) of stress-induction followed by a strong decrease after 60 min (n = 101). Error bars refer to standard error (se). F) Integrated density (IntDen) comparison between monitored proteins over time show increasing tendencies over time for Ffh and FtsY. L1 increases after 30 min and decreases after 60 min. Thus IntDen is directly influenced by the area of the cells, it is plausible that IntDen show an overall increase over time compared to mgv, which is not influenced by the cellular area. Error bars refer to standard error (se).
Results for cluster analysis show that intensity measurements are less powerful to uncover antibiotic stress response in our study compared to abnormal cell grow represented by increased cellular area (see Figure 8). For L1, 6 distinct clusters are identified to which the highest mgv is close beyond 2500 mgv (shown in green). No cell beyond a mgv of 2000 has a larger cellular area than 4 µm2 (see Figure 8 L1). This possibly indicates that higher mgv refers to a non-homogenously distributed population of cells with high L1 amount. It further supports the idea that abnormal cell shape and increased cellular area over time are the relevant indicators for monitoring puromycin induced stress response (see Figure 4, Figure 5, Figure 6, Figure 7). In contrast to L1, Ffh shows 10 clusters and FtsY 3 clusters. L1 (red), Ffh (red) and FtsY (green) show one central cluster. However, key finding of the cluster analysis is that cells of respective high protein amount have no enlarged cellular area beyond 4 µm2 for all investigated proteins. It confirms that stressed cells (indicated by enlarged cellular area) do not correspond to increased intensity measurements (compare to Figure 7E and F).
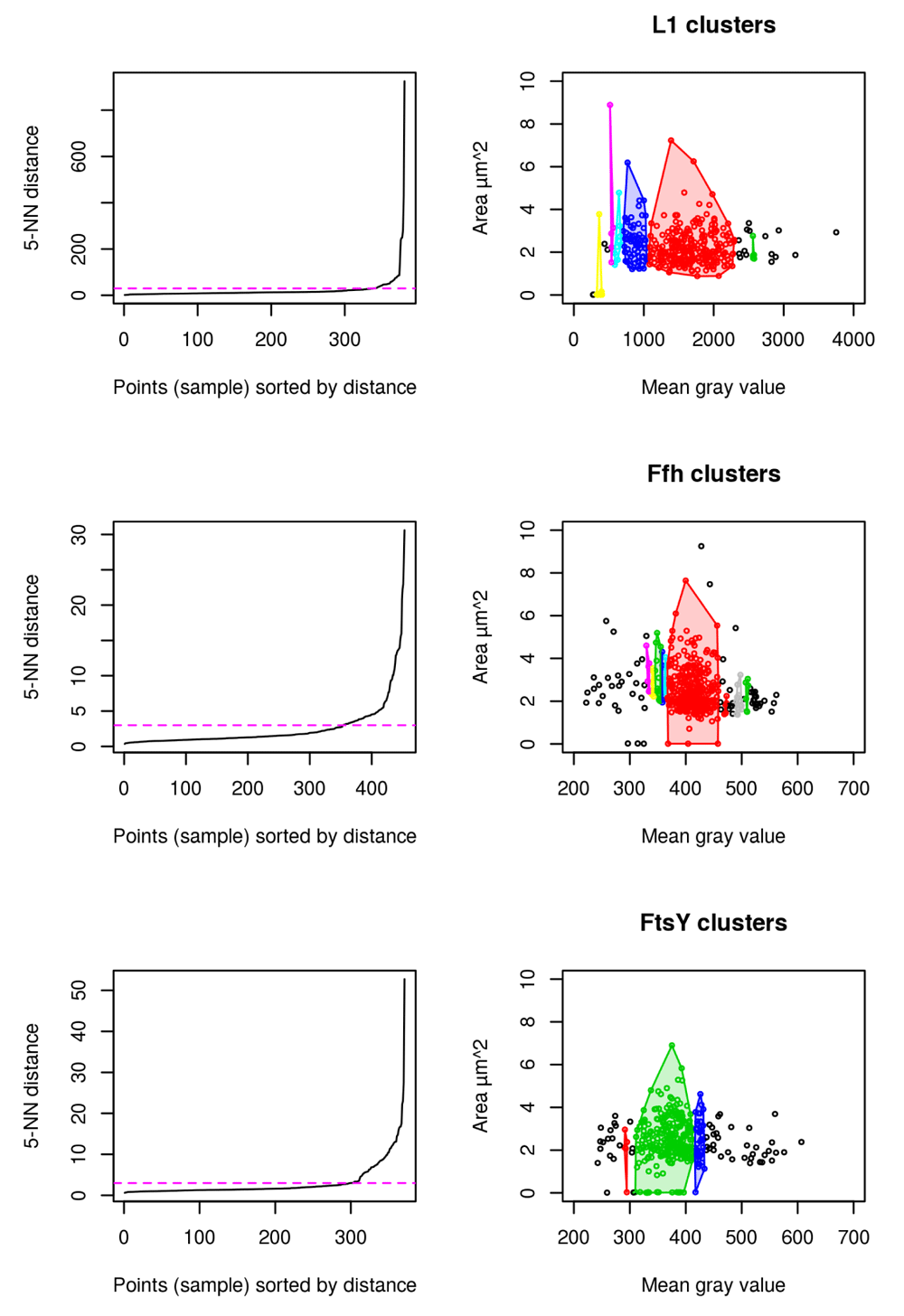
L1 (n = 381), Ffh (n = 453) and FtsY (n = 371) shows that the cells with largest area do not belong to the cells with the highest amount of protein. 5-Nearest Neighbors (NN) distance plots are used to define the appropriate epsilon values for dbscan and are adjusted manually according to knee of the curve (magenta). Resulting clusters are color coded for discrimination respectively (right).
The A.D.I.C.T. workflow is sensitive enough to monitor time-course drug-screening experiments in a reproducible manner. Binary images are very robust regarding their information content and useful for addressing complex questions involving cell shape modelling at nanoscale levels. However, binary images processed in this study are based on brightfield images and extended manual correction is necessary (see Figure 1). By using more powerful techniques like phase-contrast or specific membrane stains for instance, cell detection would be improved and the effort to manually correct binary images could be decreased further resulting in almost fully automated cell detection. Nevertheless, although we applied very basic image acquisition techniques, it is clearly demonstrated that cell division specific events can be monitored, processed and analyzed using the presented workflow. The advantage of the workflow is that every step can be redeployed and improved starting from the raw data processing to final statistical evaluation using high-level or low-level programming languages. Bias can be reduced by re-deployment and refinement of the database which enhances reproducible dissection and analysis of complex data sets in an automated fashion. The amount of useful information gathered from deploying the A.D.I.C.T. workflow in case of puromycin stress on our model organism is convincing but far away from being fully covered by this article. To sum up, our approach illustrates how powerful very basic imaging techniques can be, if applied with a robust, combined workflow and we hope that it empowers other researchers to take advantage from it for their own research tasks.
Open Science Framework: Test data for A.D.I.C.T. workflow, https://osf.io/ynkz3/46
This project contains the following files:
Binary images
Projections
ROI
merged results tables (.csv)
brightfield raw images
time-lapse raw recordings
Data are available under the terms of the Creative Commons Attribution 4.0 International license (CC-BY 4.0).
Scripts from this study are available at Github: https://github.com/Image-processing-and-analysis-workflows/A.D.I.C.T.
Archived scripts as at time of publication: https://doi.org/10.5281/zenodo.534292347.
License: GNU GPL 3.0
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)