Keywords
C-type lectin, gene encoding, LvCTL genes, white leg shrimp.
This article is included in the Genomics and Genetics gateway.
C-type lectin, gene encoding, LvCTL genes, white leg shrimp.
We would like to change in first author and add more one institution, where PhD thesis has been done by the author. Other minor changes were implemented throughout.
See the authors' detailed response to the review by Mengqiang Wang
See the authors' detailed response to the review by Hieu Tran-Van
C-type lectins (CTLs) are proteins/glycoproteins capable of reversibly binding to carbohydrates by non-covalent bonds without altering their structure (Zhang et al., 2009). Lectins can bind to cells with glycoproteins or microbial surface glycolipids and are considered pattern recognition receptors (PRR) recognizing and releasing invading microorganisms in the system. The shrimp lectin protein family is very rich and diverse; members of the eight PRR families have been identified in several shrimp species, based on structure and specificity to different sugars of the carbohydrate recognition domain (CRD). These families are lipopolysaccharide and b-1,3-glucan binding proteins (LGBPs), CTLs, galectin, thioester-containing proteins (TEP), fibrinogen-related proteins (FREP), scavenger receptors (SR), down syndrome cell adhesion molecules (DSCAM) and Toll-like receptors (TLR) (Wang and Wang, 2013). According to form and function, lectins are classified as Calnexin, C-, L-, P-, I-, R-, and S-type lens (Janeway and Medzhitov, 2002; Zhang et al., 2009). Among the lectins, CTL is the most diverse and well-studied (Zhang et al., 2009).
CTLs are a group of proteins that play an essential role in many animal biological processes, including cell signaling and pathogen recognition. CTLs are a superfamily of more than 1,000 proteins identified by having one or more well-characterized C-type lectin-like domains (CTLDs). They are divided into 17 subgroups based on their species and domain organization (Weis et al., 1998; Zelensky and Gready, 2005). CTL also plays an essential role in mammalian innate immunity. The mannose-binding lectin (MBL) is primarily expressed in the liver and released into the plasma, where it recognizes potentially pathogenic bacteria and binds to microbial surfaces. Animals can also activate the complement pathway (Thiel and Gadjeva, 2009). Similarly, CTLs are also important immune recognition receptors in invertebrates. CTLs in shrimp can currently be divided into three subgroups based on domain composition and organization: those containing only one C-type lectin domain (CTLD), those having two CTLDs, and those containing one CTLD and another part (Wang and Wang, 2013). Among them, the group containing only one CTLD was more common than the few CTLs in shrimp containing two CTLDs. The presence of two or more binding sites for each lectin molecule allows it to bind to many different cell types. The prevalence of CTLD in invertebrates is much higher than in vertebrates. Typically, CTLD is the seventh most common domain in the Caenorhabditis elegans genome, but only the 43rd most common domain in the human genome (Zelensky and Gready, 2005). Shrimp CTLD can bind multiple ligands and constructs inducible expression of CTLs. Recent studies have revealed multiple functions and mechanisms of shrimp CTLs in antimicrobial and antiviral immunity. These protective functions of CTLs in the shrimp species Fenneropenaeus chinensis are based on their ability to recognize and bind CTLDs (Sun et al., 2008). In the framework of this study, we present the results of cloning and characterization of the LvCTL genes isolated from white-leg shrimp (Litopenaeus vannamei) in Thua Thien Hue as a prerequisite for the production of recombinant LvCTL proteins for further research.
Animal use in this study was allowed by Hue University of Animal Ethics Committee with certificate reference number: HUVN0018, April 10th, 2022. Principal Researcher: Nguyen Quang Linh. All efforts were undertaken to minimize the suffering of animals.
White-leg shrimp (L. vannamei) with a weight of ~20 g per head, which were collected from shrimp ponds; the samples were aseptically dissected to collect the hepatopancreas shrimp samples and stored in liquid nitrogen, and used directly for RNA extraction.
Total RNA from hepatopancreas samples was extracted using the Gene JET RNA Purification Kit (Thermo Scientific, USA) according to the manufacturer's instructions. Electrophoresis was conducted to check the total RNA obtained on 1% agarose gel with TAE 1X at 70V for 30 min by using Power Pac 3000 (Bio-Rad). The cDNA biosynthesis was performed using Revert Aid First Strand cDNA Synthesis kit (Thermo Fisher Scientific, USA), and the obtained product was used directly for PCR amplification.
The sequences of LvCTL encoding genes were obtained from GenBank to design a specific primer (Table 1). PCR was performed using the following conditions: 95°C for 2 min, and then 40 cycles of 95°C for 1 min, 55°C for 30 s, and 72°C for 1 min, followed by a final extension at 72°C for 7 min. PCR products were confirmed by electrophoresis at 70V for 35 min on 1% agarose gel with TAE 1X stained SafeViewTM Classic (abm, Canada).
| Genes | Primers | Sequences (5’-3’) | Product sizes | References |
|---|---|---|---|---|
| LvCTLD | LvCTLD-F/LvCTLD-R | GACTGCACGGGCGACGAG | 888 | (Junkunlo et al., 2012) |
| TTAGACAGCGGTGACGCAGAG | ||||
| LvCTL3 | LvCTL3-F/LvCTL3-R | CTCTCCAATCCCATCTCAATC | 444 | (Li et al., 2014) |
| CTATTTCTCACAGATAATG | ||||
| LvCTL4 | LvCTL4-F/LvCTL4-R | TGTTCTGATGGCTGGCTGG | 417 | (Li et al., 2015) |
| TTAAACCATACAAATGGGATG | ||||
| LvAV | LvAV-F/LvAV-R | ACCTTATACGAGAAAAGTG | 474 | (He et al., 2015) |
| TCAAAACTTGTCAGCAGATG | ||||
| LvLdlrCTL | LvLdlrCTL-F/LvLdlrCTL-R | GAGTGTACCAACAGGGAC | 855 | (Liang et al., 2019) |
| TCACGCCCTCTCACTGGG |
| Characteristics | LvCTL3 | LvCTL4 | ||
|---|---|---|---|---|
| Li et al. (2014) | This study | Li et al. (2015) | This study | |
| Gene full lenghts (bp) | 579 | - | 563 | - |
| Coding segment (bp) | 492 | 444 | 471 | 417 |
| Polypeptide chain (amino acid) | 163 | 147 | 156 | 138 |
| Molecular weight (kDa) | 17.9 | 16.91 | - | 15.75 |
| Isoelectric point (pI) | 4.80 | 4.66 | - | 4.58 |
| Domain | 1 | 1 | - | 1 |
| α helix | - | 2 | - | 2 |
| β strain | - | 9 | - | 8 |
PCR products were recovered from agarose gels and purified using the GeneJET Gel Extraction kit (Thermo Scientific, USA). Purified PCR products were cloned into pGEM T-easy vector with T4 DNA Ligase for 1 hour at room temperature (Promega, USA), then, the recombinant vectors were transferred into Escherichia coli TOP10 bacteria using the heat shock method. The recombinant vectors were selected on LB medium (Luria Bertani Agar, Merck, Germany) supplemented with 50 μg/mL Amp, X-Gal/IPTG. E. coli cells carrying the recombinant vector (white colonies) were then inoculated with 5 mL of liquid LB medium supplemented with 50 μg/mL Amp, and DNA plasmids were isolated using the GeneJET Plasmid Miniprep Kit (Thermo Fisher Scientific, USA). The isolated DNA plasmid was examined by 1.0% agarose gel electrophoresis at 70V for 35 min and used for LvCTL sequencing (Ha et al., 2018). The sequence of the LvCTL genes was analyzed using the Sanger method (First base Company, Malaysia). The results were checked using BioEdit software and then compared with those published in GenBank using the BLAST tool. A phylogenetic tree of CTL encoding genes was built using MEGA 11 software with the Neighbor-Joining algorithm (Tamura et al., 2021).
Characterization of LvCTL proteins was determined using bioinformatics tools, including amino acid sequence translation with Expasy, protein domain using SMART (Letunic et al., 2021), spatial structure model using Phyre2 (Kelley et al., 2015), isoelectric point using IPC2 (Li et al., 2014).
After PCR with specific primers, the results showed that two DNA fragments were amplified and expressed approximately at 450 and 420 bp in size, respectively. According to the theoretical length of the cDNA fragment of the LvCTL3 and LvCTL4 genes. PCR reaction using specific primers with pGEM vector as template was performed, electrophoresis images showed very specific PCR products (Figure 1), which proved that LvCTL genes was successfully attached to pGEM vector T-easy (referred to as the recombinant vector pGEM/LvCTL3 and pGEM/LvCTL4). Therefore, the recombinant vectors were used to analyze the nucleotide sequences. The previous reports indicate several CTLs in some shrimp species, such as: Litopenaeus vannamei, L. schmitti, L. setiferus, Fenneropenaeus chinensis, F. merguiensis, Penaeus monodon, Marsupenaeus japonicus and Macrobrachium rosenbergii (Wang and Wang, 2013). We constructed a phylogenetic tree from the LvCTL3 and LvCTL4 deduced amino acid sequences that isolated from genes compared with the published reference sequences of other shrimp species. The results show that LvCTL3 and LvCTL4 deduced amino acid sequence belong to two groups in the dendrogram and our arrangements were similar to the published of white-leg shrimp (L. vannamei) but different from other shrimp or crustacean species (Figure 6).

M: DNA ladder (GeneRulerTM 1 kb DNA Ladder, Thermo Scientific, USA). NC: Negative control. 1-5: PCR product with cDNA of LvCTL3-LvCTL4-LvAV-LvCTLD-LvLdlrCTL genes. 6-7: PCR product of LvCTL3 gene from 2 different colonies. 8-9: PCR product of LvCTL4 gene from 2 different colonies.
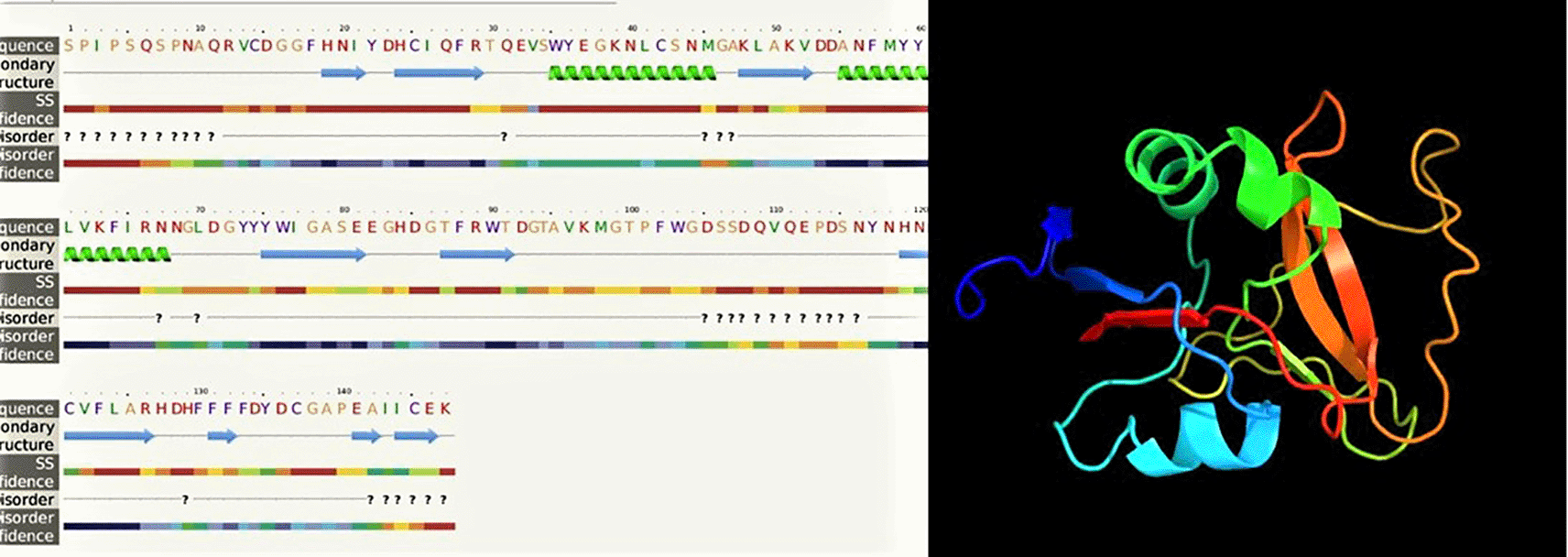
In our study, the nucleotide sequence of LvCTL3 PCR product was 444 bp in length, encoding a polypeptide sequence of 147 amino acids. The nucleotide sequence of LvCTL3 PCR product was 417 bp in length, encoding a polypeptide sequence of 138 amino acids. These were fragment of genes containing the open reading frame (ORF) region cut off the peptide signal, intended to be used to produce recombinant LvCTL protein for other studies related to recombinant protein production. For the conserved domain determined using the SMART program, the ORF sequence of LvCTL3 had one domain (CTLD) from amino acid position 14 to 146. LvCTL3 had a molecular weight of 16.91 kDa, and the isoelectric point (pI) was 4.66. The results of spatial structure prediction showed that the LvCTL3 molecule had two alpha helices (16%) and nine beta strands (28%) (Figure 7). Meanwhile, the ORF sequence of LvCTL4 had one domain (amino acid 1 to 137), a molecular weight of 15.75 kDa and a pI was 4.58. LvCTL4 structure had two alpha helices (17%) and eight beta strands (32%) (Figure 8).
The results of sequence analysis showed a change in the isolated LvCTL genes compared with the published genes. The isolated LvCTL3 fragment was 444 bp in size (including the stop codon), 98.87% homologous to the published gene (439/444 nucleotides compared with KF156943) (Figure 2). The change of the LvCTL3 gene resulted in a difference in the sequence of the deduced amino acid sequence, the highest similarity obtained was 97.28% (143/147 amino acids) compared with the reference sequence AGV68681. Meanwhile, the LvCTL4 gene fragment was 417 bp, 99.52% homologous to the gene of KM387560 (415/417 nucleotides) (Figure 3). However, the change of the LvCTL4 gene resulted in no change in the sequence of the deduced amino acid sequence (100% similarity) compared with the reference sequence (AKA64754).






According to Li et al. (2014), the LvCTL3 gene from shrimp (L. vannamei) has a length of 579 bp, in which the coding segment is 492 bp (from nucleotide positions 25 to 516), encoding a polypeptide chain with a length of 163 amino acids, and has an estimated molecular weight of 17.9 kDa, an estimated pI of 4.80 (Li et al., 2014). The LvCTL4 gene, also from L. vannamei shrimp, has a length of 563 bp, in which the coding segment is 471 bp (from nucleotide positions 27 to 497). The ORF encoded a protein of 156 amino acids consisting of a single CTLD (residues 19-155) and a putative signal peptide (residues 1-18). Tissue expression analysis showed LvCTL4 was distributed with high levels in the gills, intestine, epithelium and hepatopancreas (Li et al., 2015). Meanwhile, LvTRAF3 was also cloned and characterized from the shrimp L.vannamei; it had a transcript of 3,865 bp, with an ORF of 1,002 bp, and encoded a polypeptide of 333 amino acids, with a calculated molecular weight of 38.6 kDa (Li et al., 2020).TRAF3 functions as a regulator of innate immune response that involves many cellular processes.
When comparing to the whiteleg shrimp (L. vannamei) deduced amino acid sequence from LvCTL3 genes, the resulted in 97.28% similarity to the reference sequence of AGV68681 or 96.60% to XP_027212325, while LvCTL4 deduced amino acid sequence was 100% similar to the Genbank reference sequence of AKA64754 and XP_027228060. The LvCTL3 and LvCTL4 deduced amino acid sequence were 60.69% (XP_042883674) and 80.43% (QEX50549) homologous to kuruma prawn (Penaeus japonicus), respectively. Compared with black tiger shrimp (Penaeus monodon), the LvCTL3 deduced amino acid sequence was 59.18% similar to the reference sequence by XP_037787306 while LvCTL4 was 88.21% similar to that of XP_037800826. Compared with other crustacean species, the difference in LvCTL3 and LvCTL4 sequences was higher. The similarity of the LvCTL3 deduced amino acid sequence was only 45.58% compare to the AYD41573 reference sequence from red swamp crayfish (Procambarus clarkii) or 40.25 % to the XP_042233235 reference sequence from the American lobster (Homarus americanus). Meanwhile, LvCTL4 amino acid sequence was only 59.12% similar to the XP_045611199 reference sequence from red swamp crayfish, 60.14% to XP_042237721 from the American lobster, or 57.258% to the XP_045118055 sequence from gazami crab (Portunus trituberculatus).
The amino acid sequence alignments of two proteins (Figure 4 & Figure 5) that may cause protein secondary structure shifts (Li et al., 2015). The form of a protein is related to its function. Knowledge of protein’s 3D structure is a huge hint for understanding how protein’s work. Although other refinement tools such as homology-modeling tools based on high sequence similiritues provide higher quality structure. However, our 3D structures have been modelled with 99.9% and 99.8% confidence by the single highest scoring template with the 3D structure of LvCTL3 and LvCTL4, respectively. This showed that the function of these proteins is not changed. Protein folding is determined by the physicochemical properties that are encoded in the amino acids. Although most of the predictions were highly accurate, the system that we used is not perfect. Programs such as Alpha Fold will exponentially increase our general understanding of different biological processes. So the 3D structure of these proteins should be further study by another program to provide higher-quality structures. And the simulation of these proteins at pH of shrimp stomach (5.7) should be considered for further experiments.
In this study, two LvCTL3 and LvCTL4 genes from white-leg shrimp were successfully isolated, cloned, and sequenced. The ORF of LvCTL deduced amino acid sequences from gene had homology levels of 98.87% (LvCTL3, KF156943) and 99.52% (LvCTL4, KM387560 respectively) compared to the published reference sequence. The predicted LvCTL proteins have one conserved domain and are defined as C-type lectins. They can be used for heterologous expression and scale-up production of recombinant C-type lectin in the future to add to the aquatic feed to enhance immunity and prevent disease in shrimp.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)