Keywords
Secretome, Mesenchymal stem cells, Immunomodulator, In vivo, Safety
Secretome, Mesenchymal stem cells, Immunomodulator, In vivo, Safety
Version 2 is a revised version of Version 1, reflecting the reviewers' suggestions and questions. The main revisions include the correction of author affiliations, as it was identified that the affiliations in Version 1 were incorrect. Other revisions have been made in the abstract, introduction, methods, results, and discussion sections. Specifically, the reviewers raised concerns regarding the rationale for conducting the study in healthy animals and pointed out the need for clarification regarding references that mixed stem cells with the investigated product, which is secretome. Consequently, we have revised the introduction and discussion accordingly. Additionally, the methods and results sections were updated in response to the reviewers’ requests for new data and further explanation regarding the previous methodology, including the justification for using the intramuscular route instead of the more commonly used intravenous route. Figures (3 and 6) and Tables were updated to reflect these changes.
See the authors' detailed response to the review by Francesca Pischiutta
See the authors' detailed response to the review by Basak Isildar
The immune system is a sophisticated network designed to protect the body from harmful pathogens. It is generally divided into two main categories: the innate immune system and the adaptive immune system.1 The innate immune system serves as the first line of defence mechanism against pathogens, using processes like phagocytosis to eliminate harmful invaders before they enter the bloodstream and cause further complications.2 Macrophages in the tissue remove pathogens or foreign particles that enter the tissue, while the spleen helps remove pathogens or foreign particles that have entered the bloodstream by filtering the blood and facilitating their destruction.3
Maintaining homeostasis in the immune system is crucial to ensure that it is prepared to protect the body whenever pathogens or other harmful substances enter. Factors such as daily stress, lifestyle choices, and environmental hazards can weaken the immune system and increase susceptibility to illness. Therefore, it is essential to maintain and enhance immune function to prevent infections and other diseases.4,5
Researchers are finding that the secreted materials from mesenchymal stem cells (MSCs), known as the “secretome,” and the liquid containing these materials, referred to as “conditioned medium,” are promising as immunomodulatory agents.6 The secretome includes soluble factors like immunomodulatory molecules, cytokines, and growth factors, as well as vesicular factors such as extracellular vesicles (EVs) including exosome.7,8 Notably, MSC-derived exosomes, which are small vesicles containing proteins, nucleic acids (like mRNA and miRNA), and lipids, have shown significant potential.9
Overexpression of IL-10 in the secretome of human umbilical cord MSCs can enhance macrophage function,10 while MSC-derived exosomes can improve the anti-inflammatory properties of macrophages and aid in reducing inflammation.11 Additionally, apoptotic bodies from MSCs can influence macrophage behavior in wound healing models, and MSC exosomes have the potential to inhibit the proliferation, activation, and cytotoxicity of natural killer cells.12 Furthermore, miRNAs present in MSC-derived EVs play a crucial role in regulating macrophage functions during inflammation.13,14
Although the immunomodulatory effects of the secretome are well-documented, there is limited information on how secretomes derived from human umbilical cord MSCs (UC-MSCs) support immune function in healthy or normal conditions. This study aims to address this gap by evaluating several key aspects of the UC-MSC secretome in a healthy context. We measured interleukin-10 (IL-10) to understand its role in phagocytosis, profiled the miRNA content to uncover the mechanisms behind the secretome’s effects, characterized the EVs to explore their therapeutic potential, and assessed the impact of the secretome on phagocytosis in vivo. Additionally, we conducted an acute toxicity assay to ensure the safety of UC-MSC-derived secretomes.
The experimental protocol was approved by the Medical and Health Research Ethics Committee (MHREC) Faculty of Medicine Universitas Gadjah Mada (KE/0108/01/2022).
PT Tristem Medika Indonesia provided the MSC secretome. The MSC secretome was produced in four batches, on 17th September 2021 (Batch I), 27th January 2022 (Batch II), 10th April 2022 (Batch III), and 5th July 2022 (Batch IV), respectively. The MSC secretome production procedure followed the PT Tristem Medika Indonesia method. Briefly, the umbilical cord tissue was collected from eligible consenting donors following caesarean delivery and transported to the laboratory using a sterile bottle with DPBS (Gibco) with 2% penicillin-streptomycin (Sigma Aldrich, Germany). The tissue was rinsed and cleaned from blood using a similar solution that was used for transport. The blood vessel was removed and cut transversally into 1-cm segments. Then, it was cut longitudinally into halves. The tissues were rinsed using DPBS (Gibco, Thermo Fisher Scientific, USA), minced into 2-5 mm small pieces, and explanted to a 100 mm cell dish (Nunc, Thermo Fisher Sceintific, USA) in MEM Alpha Glutamax (Gibco) supplemented with 10% UltraGro (AventaCell Biomedical, USA) and 1% penicillin-streptomycin. The explants were incubated at 37°C, 5% CO2 and 2-8% O2. The medium was replaced every four days, and the tissues were removed once the cells were migrated. After 14 days, the cells were harvested and subcultured using the antibiotic-free complete medium with a seeding density of 8000 cells/cm2. The UCMSCs were further expanded until passage four to create a two-tier cell bank, master cell bank (passage two), and working cell bank (passage four). The working cell banks were thawed and cultured to passage six. After cells reached 90% confluency (day three), the culture medium was replaced with MEM Alpha without phenol red (Gibco) and further cultured for another three days. After three days, the conditioned medium was harvested, filtered using a 0.22 μm membrane (Nunc), and filled into cryovials (Corning) with 1.5 mL volume. The secretome products were stored at -20°C. The secretome was stored at -80°C for long-term preservation. However, for short-term storage, such as when preparing for studies, we stored the secretome at -20°C. The characterization of MSCs was performed using flow cytometry (FACS) at the commercial lab testing facility, PharmaMetricLabs®. Several parameters were measured, including CD105, CD90, CD73, CD45, CD34, CD14, HLA-DR, and CD19.
Standard ELISA procedures using EliKineTM Human IL-10 ELISA KIT (Abbkine, China) were used to evaluate the IL-10 concentration in the secretome. The measurements of IL-10 in secretome were conducted based on the protocol provided by the kit manufacturer. The absorbance was read at 450 nm using an iMar Microplate reader (Bio-Rad, UK). The ELISA data were analyzed using Microplate Manager Software (Bio-Rad).
Exosomes were isolated from MSC secretome using Total Exosome Isolation from biofluids (Invitrogen, Thermo Fisher Scientific, USA). The starting volume of the secretome used was 10 mL and prepared according to the manufacturer’s instructions. The isolated exosomes were then diluted using PBS as much as 1 mL. Diluted exosomes were stored in -80°C for further characterization.
The size distribution and concentration of isolated exosomes were examined using NTA. The NTA (ViewSizer 3000 – Horiba) instrumentation was equipped with multi-Laser; Blue-445 nm, Green-520 nm, and Red-635 nm, and a fluorescent detector with 450, 550, and 650 nm filters so that analysis could be done using a fluorescent marker. The volume required for the measurement process was as much as 500 μL exosome in PBS, and the observed particle sizes were limited to a 30 to 150 nm range.
For TEM analysis, as much as 200 μL of isolated exosomes were diluted using PBS to reach a volume of 1 mL exosome in PBS. Exosomes were dripped onto the 400 mesh Cu coated grid, allowed to absorb, and dried using filter paper. Negative staining was applied based on protocol by Jung and Mun, 2018,15 using 1% uranyl acetate. The negative staining was dripped on the grid and allowed to be absorbed, then dried with filter paper. After that, the grid stored in an EM grid box for TEM observation. TEM visualizations were performed using JEOL 1400 Transmission Electron Microscope at 120 kV, and images were captured using a fully integrated high resolution camera system (8 million pixel CCD camera). Exosome particle size measurements were carried out using ImageJ software version 1.53K.
The miRNA profiling started by isolating the exosome using exoRNeasy Midi Kit (Qiagen) based on the manufacturer’s protocols. The total RNA calculation was done using a NanoDrop (Thermo Scientific) and the miRNA profiling was conducted using the Nanostring OID NGS-036 (NanoString Technology, USA). Then, the miRNA data analysis was conducted using nSolver Analysis Software 4.0, with which the ten highest normalized expressions of the miRNA were obtained. The endogenous housekeeping gene was beta-actin. The threshold for reading background subtraction was determined using negative control read.
The prediction of the top 10 miRNA biological activity was done using free software DIANA miRPath v3.0 tools available From DIANA Lab, supported by experimental interaction from DNA-TarBase v.7.0. TarBase v7.0 is a combination of half a million miRNAs derived from gene interactions from hundreds of publications and more than 150 CLIP-Seq libraries.16
Twenty male Swiss albino mice, 6-8 weeks old, weighing 15 to 25 g, were used in the study. The animal’s room temperature is set to 22° ± 3°C, with a relative humidity of 30–70%, 12 hours of light, and 12 hours of darkness. Animals are fed according to laboratory standards and given indefinitely (ad libitum). The animals were maintained under standard conditions according to Regulation of the Head of National Agency of Drug and Food Control (BPOM) No. 7 of 2014 concerning guidelines for in vivo tests.16 The person conducting the animal handling was not blinded to the treatment group. However, the person conducting the measurement of the carbon clearance assay was blinded to the treatment group.
After the test was complete, all experimental animals were sacrificed using ketamine at a dose of 80 mg/kg BW. Ketamine is injected intraperitoneally. The experimental protocol was approved by the Medical and Health Research Ethics Committee (MHREC) Faculty of Medicine Universitas Gadjah Mada (KE/0108/01/2022).
Twenty healthy male mice were randomly divided into four groups. Each group consisted of five mice, and all received an intramuscular injection of secretome or MEM-α basal medium at 0.025 mL in each thigh. The first group received 0.00625 mL of secretome intramuscularly in each thigh (dose 1). The second group received 0.0125 mL of secretome intramuscularly in each thigh (dose 2). The third group received 0.025 mL of secretome intramuscularly in each thigh (dose 3). The control group received 0.025 mL of sterile MEM-α basal medium in each thigh (negative control). After three days of treatment, all the groups received 0.1 mL of carbon ink suspension through the tail vein. Blood was collected at an interval of 0 and 15 min immediately after injection of carbon suspension. Measurement of carbon in the blood was determined using a spectrophotometer at 675 nm. The phagocytic index was calculated based on optical densities value with the following formula.
Note:
OD1 is the optical density at 0 minutes after carbon injection
OD2 is the optical density at 15 minutes after carbon clearance injection
t2 is 15 minutes
t1 is 0 minutes
Total mRNA was extracted from the spleen using the FavorPrep™ Tissue Total RNA Mini Kit (Favorgen) according to the instructions provided in the kit. The extracted mRNA was then converted into complementary DNA (cDNA) using the ReverTraAce™ qPCR RT Master Mix (Toyobo). The mRNA expression was evaluated using the RT-qPCR SensiFAST™ SYBR® Low-ROX Kit (Bioline), with Beta Actin as the housekeeping gene. The beta actin primers used were: F 5′-TCT CCA CCT TCC AGC AGA TGT-3′ R 5′-GCT CAG TAA CAG TCC GCC TAGA-3′. The CD68 primers were: F 5′- CGC AGA CAA TCA ACC TA-3′ R 5′-GGC ATG GTG AAG TGA GG-3′.
Female Swiss albino mice were used in this test. Healthy female mice at 8-10 weeks old were used because, generally, female animals are more sensitive to toxic substances than male animals.16 The method for conducting the acute toxicity test was modified fixed dose according to Regulation of the Head of National Agency of Drug and Food Control (BPOM) No. 7 of 2014.16 There are two parts to the fixed-dose methods: preliminary and main parts. In this preliminary part, we used two mice divided into two groups, the treatment group, and the control group. The mice received secretome or MEM-α basal medium intramuscularly with the highest possible volume that can be given intramuscularly to mice, which was 0.05 mL in each thigh or 0.1 mL per mouse. The mice were randomly assigned to receive treatment or control. Observations of their behavior and clinical signs were conducted in the first 30 minutes and every 4 hours for 24 hours. After 24 hours, the mice were observed daily until day 15. Since there was no mortality in the preliminary parts of the acute toxicity test, the main test was carried out with the same dose as the dose used in the preliminary part. In the main test, we used four mice in the secretome group and one mouse in the control group. The mice were observed in the first 30 minutes after injection, every 4 hours during 24 hours, and every 24 hours until day 15. On day 15, all mice were killed, and all vital organs were examined.
The behavioral examination includes the following assessments: skin condition, fur piloerection, corneal reflex and corneal opacity, mucous membrane color, type and rate of respiration, palpebral reflexes, digit reflexes, paralysis, seizures, torticollis, somatomotor activity, reverse walking, tremors, salivation, sleep patterns, defecation, and urination.
The characterization of MSCs was performed using flow cytometry (FACS). The complete results are presented in Table 1.
The IL-10 concentration measured by ELISA showed that the concentration of IL-10 was 0.272-1.951 pg/mL (Figure 1).
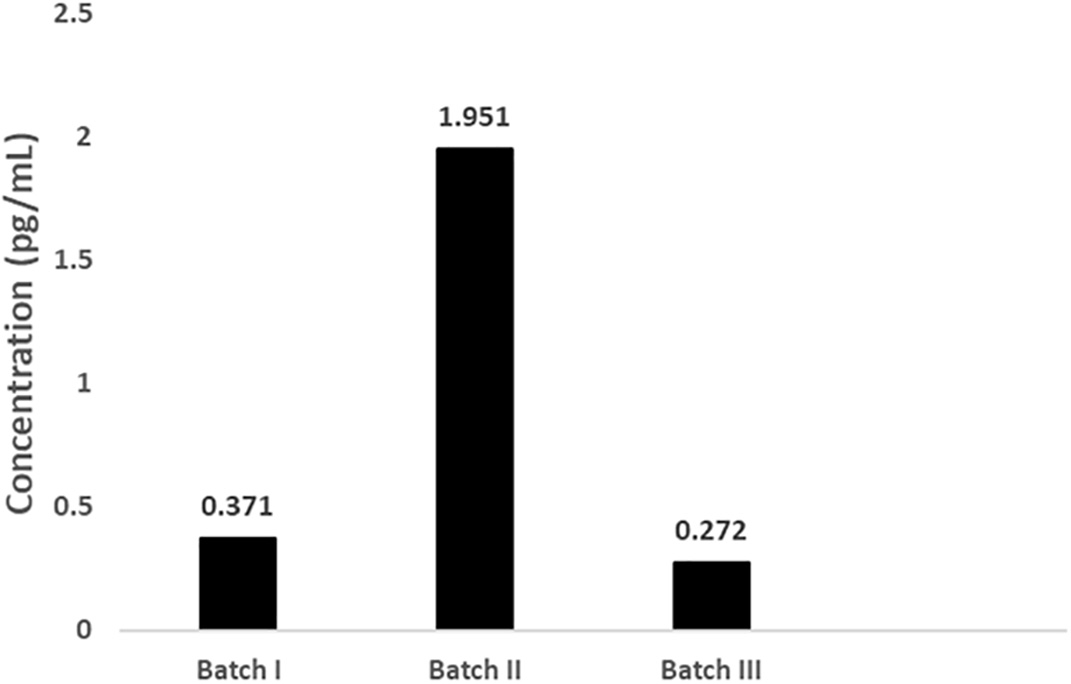
The concentration of IL-10 in the secretome varied from 0.272 to 1.95 pg/mL.
The mean particle size in secretome and water was 117 nm and 104 nm, respectively. The particles in size 30-150 nm in secretome and water were 3.1×108 and 1.5×106, respectively. Figure 2 presents the particle examination result using NTA.
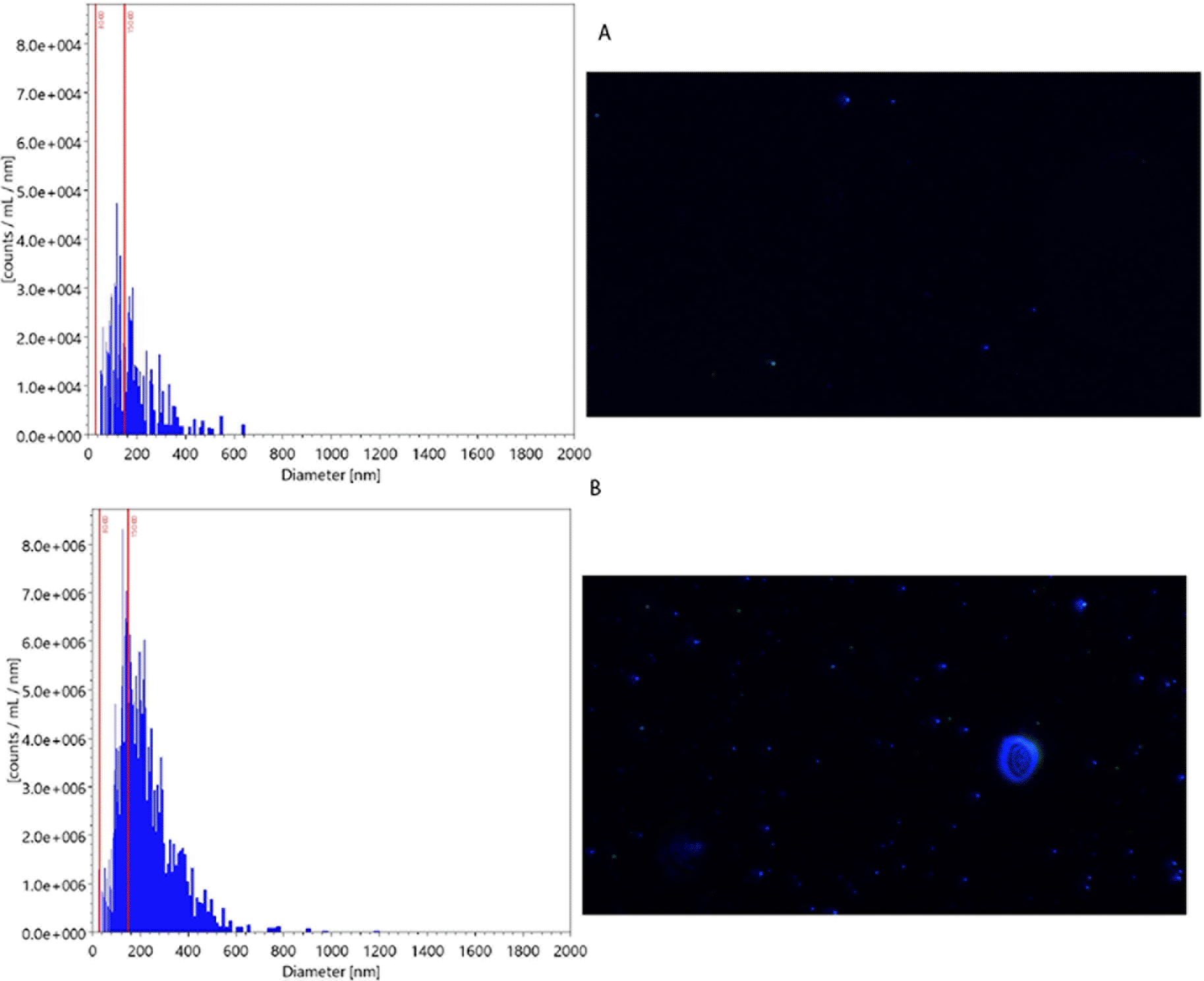
The graph on the left side shows the particle distribution across the different sizes. The picture on the right side shows the particle picture in measured samples. The mean particle size (x-axis) in the secretome was larger than those in water (117 nm (B) vs104 nm (A), respectively). The count of particles (y-axis) in size 30-150 nm was also greater in secretome than in water (3.1×108 (B) vs 1.5×106 (A), respectively).
The visualization using TEM revealed that exosomes had been successfully isolated within the expected size range (30-100 nm). The analyzed particles’ morphology (Figure 3) was consistent with NTA results.
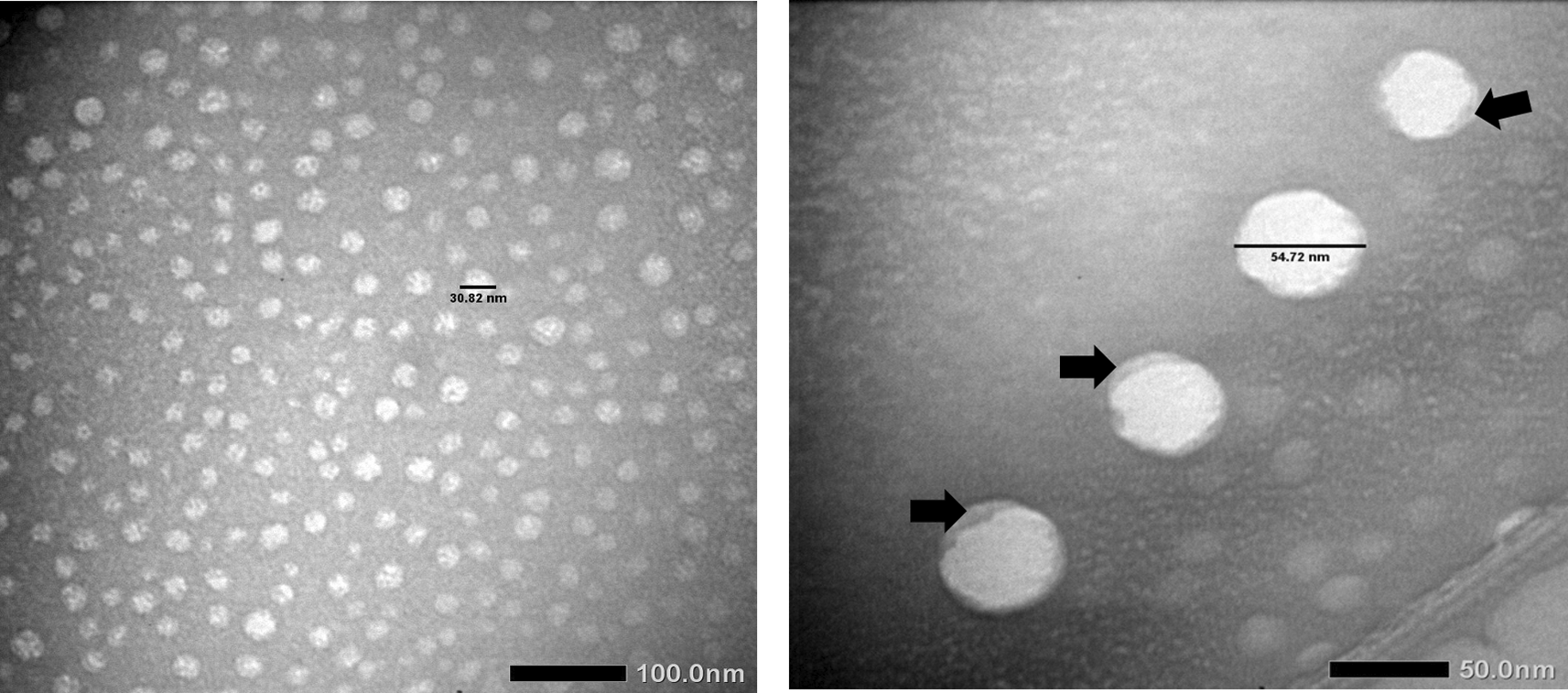
The size and morphology of the isolated exosome were varied. The size of the exosome was within the expected size range (30-100 nm). The double membrane appearance is pointed by the black arrow.
The 10 highest reads of miRNA normalized by beta actine are presented in Table 2. The 10 miRNA with highest read were hsa-miR-23a-3p, hsa-miR-4454 + miR-7975, hsa-miR-1915-3p, hsa-miR-4488, hsa-miR-125b-5p, hsa-miR-21-5p, hsa-miR-130a-3p, hsa-miR-320e, hsa-miR-494-3p, and hsa-miR-4516.
The predicted biological activity of the top 10 miRNA in the MSC are presented in Figure 4.
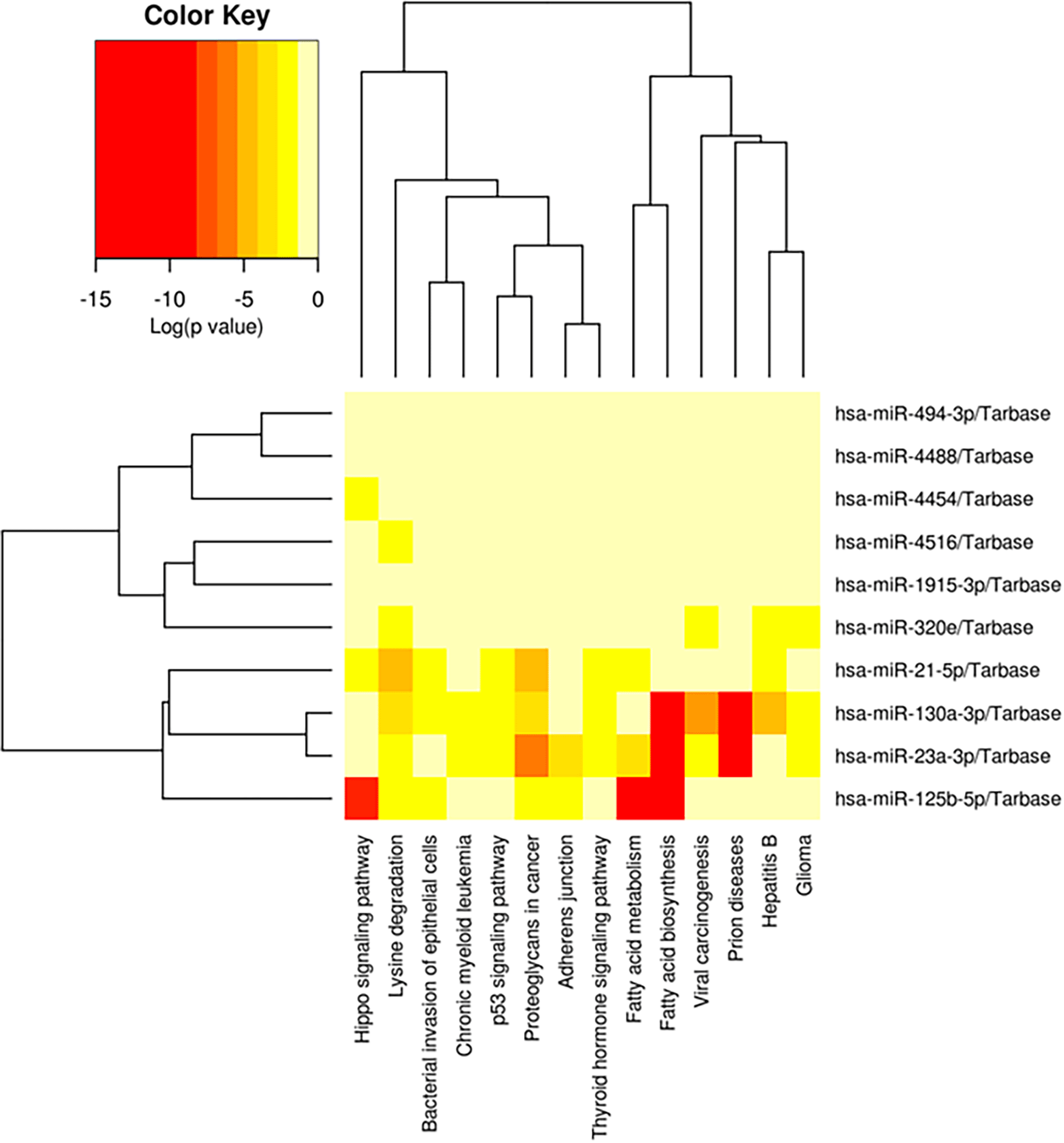
The prediction of related biological activity of the 10 highest miRNA in the secretome MSC was fatty acid biosynthesis, fatty acid metabolism, prion diseases, and hippo signalling pathway (red color).
The biological activity that were most related to the miRNA in the secretome MSCs were fatty acid biosynthesis, fatty acid metabolism, prion diseases, and hippo signalling pathway.
The phagocytic index was significantly different between those in the control and 0.025 mL group (dose 3). Figure 5 shows the phagocytic index calculation of every group.
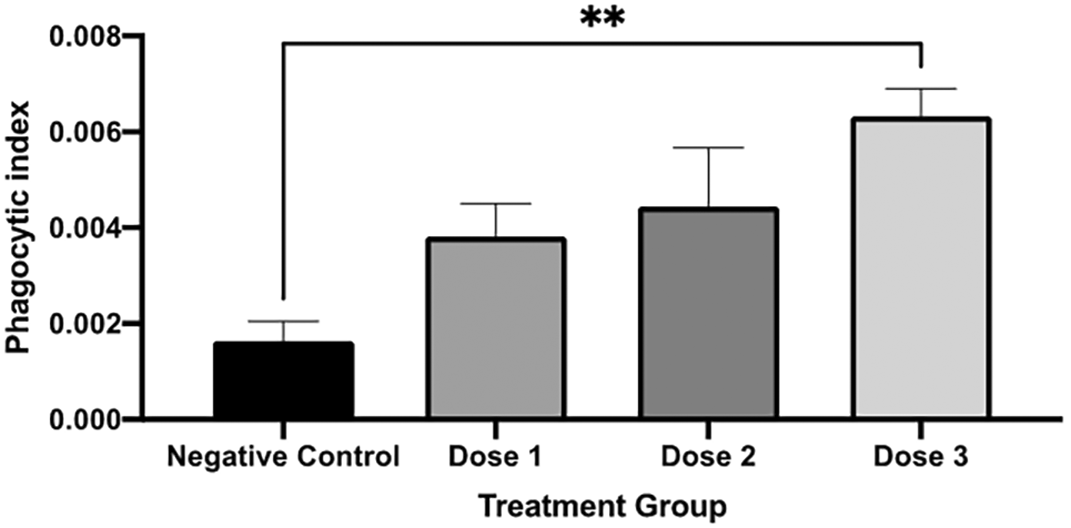
The phagocytic index was significantly higher in dose 3 compared to those in the negative control. A negative control is without secretome treatment. * p < 0.05 vs negative control.
The CD68 mRNA expression of the spleen was measured using quantitative PCR. The expression was shown in Figure 6.
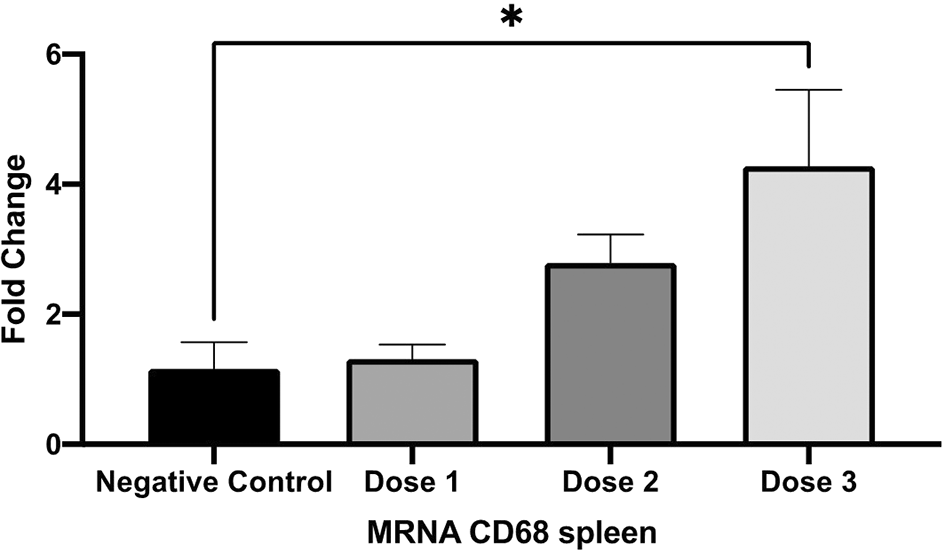
The spleen CD68 mRNA expression was significantly higher in dose 3 compare to those on negative control. A negative control is without secretome treatment. * p < 0.05 vs negative control.
The mice treated with the highest dose of secretome showed the highest mRNA CD68 expression in the spleen.
In the preliminary part of the acute toxicity test, the mice received secretome or MEM-α basal medium intramuscularly of the highest possible volume that can be given intramuscularly to mice which is 0.05 mL in each thigh or 0.1 mL per mice. There was no abnormality in the following behavioral assessment: skin condition, fur piloerection, corneal reflex and corneal opacity, mucous membrane color, type and rate of respiration, palpebral reflexes, digit reflexes, paralysis, seizures, torticollis, somatomotor activity, reverse walking, tremors, salivation, sleep patterns, defecation, and urination of the mice in the control and treatment groups. The body weight (Table 3) was also similar between the control and treatment groups. Therefore, the main study was conducted using a 0.1 mL dose per mouse.
The body weight was similar in control and treatment groups.
| Group | Body weight (g) | ||
|---|---|---|---|
| Before treatment | Day 7 | Day 15 (14 days after treatment) | |
| Control | 37.5 ± 2.1 | 38.5 ± 0.7 | 39 ± 1.4 |
| Treatment | 40.8 ± 4.9 | 41.4 ± 4.2 | 42 ± 6.4 |
The relative organ weight was measured by dividing the organ weight by the mouse’s body weight. The relative organ weight was also similar between groups (Table 4).
The relative organ weight was similar in control and treatment groups.
The secretome used in our study was derived from umbilical cord mesenchymal stem cells (MSCs). Flow cytometry analysis (FACS) was performed to confirm the identity of the cells as true MSCs. The analysis showed high expression levels of CD105, CD90, and CD73, while markers such as CD45, CD34, CD14, HLA-DR, and CD19 were present at very low percentages. According to established criteria, MSCs should express CD105, CD90, and CD73, but not CD45, CD34, CD14, HLA-DR, or CD19.17 This confirms that the cells used to produce the secretome were indeed MSCs.
The secretome also contains extracellular vesicles (EVs), which we examined using nanoparticle tracking analysis (NTA). The NTA results showed that secretomes contains 3.1 × 108 particles with diameters ranging from 30 to 150 nm, which is consistent with the typical size of exosomes which is part of EVs.18 We further examined these particles using transmission electron microscopy (TEM). The TEM results confirmed the size of the particles detected by NTA and revealed a double-membrane structure, a characteristic feature of extracellular vesicles. This double membrane is similar to the cell membrane, as extracellular vesicles are produced by cells and released outside the cells, carrying a portion of the cell's membrane.19
Our study presents the secretome effect on phagocytic activity in mice by carbon clearance assay. Phagocytosis is part of the natural immune system, which acts as a first-line defence against the intrusion of foreign matter. The injected carbon is phagocytes by reticuloendothelial systems (RES) cells which consist of phagocytic cells. This process involves opsonization and complement activation. The faster the clearance, the better the phagocytic activity.20 The phagocytic index in mice receiving the highest dose of secretome was significantly higher than those in control groups. Thus, the phagocytic activity of the mice treated with secretome was better than that of the control group.
Our study also shows that the mice treated with secretome present higher expression of mRNA CD68, the pan-macrophage marker. CD68 is a transmembrane glycoprotein expressed mostly in macrophage and mainly associated with endosomal/lysosomal compartment which make it suggested to involve in the recognition and processing of antigens or foreign bodies.21 Macrophage in the spleen play an essential role in maintaining body homeostasis by removing exogenous matter, damaged blood cells, and pathogen by protecting the body from potential harm.22
We suggest that the carbon particles injected during the carbon clearance assay serve as exogenous matter, which may induce CD68 mRNA expression as the macrophage respond to exogenous matter. Increase CD68 expression is often related with immune activation, inflammation, or tissue remodelling, reflecting the increase of macrophage activity in response to infection, chronic inflammation, or other pathologic stimuli.23 This upregulation may indicate an enhance of macrophage activation and greater capacity of phagocytosis and immune defence, which are critical for maintaining immune response.
The secretome derived from mesenchymal stem cells (MSCs) is known to contain a variety of bioactive substances, including soluble proteins, and extracellular vesicle (EVs) such as exosomes.24 Among these, cytokines are well-known for their roles in modulating the immune system.
This study measured the level of interleukin-10 (IL-10) in the secretome, focusing on its potential impact on phagocytosis. IL-10 is recognized for its role in modulating phagocytic activity, with studies in infectious models showing that IL-10 can enhance macrophage phagocytosis.25 However, our findings revealed that the IL-10 concentration in the secretome was very low, suggesting that the enhanced phagocytic activity observed in secretome-treated mice may not be due to IL-10. Additionally, the secretome was administered via intramuscular injection, which might limit the distribution of proteins to systemic circulation and target organs such as the spleen.
The secretome also contains a wide range of microRNAs (miRNAs) encapsulated within exosomes.26 In this study, we characterized the miRNA profile of the secretome and identified the top 10 miRNAs present. In silico analysis was conducted to predict the biological pathways associated with these miRNAs, revealing significant involvement in pathways such as the Hippo signaling pathway, lysine degradation, and bacterial invasion. Based on the insilico analysis, key miRNAs involved in these pathways include hsa-miR-125b-5p, hsa-miR-23a-3p, hsa-miR-130a-3p, hsa-miR-21-5p, hsa-miR-320e, hsa-miR-4516, and hsa-miR-4454.
The Hippo signaling pathway is known for its role in regulating cell proliferation, survival, and differentiation, as well as maintaining immune system homeostasis and modulating macrophage-related inflammatory responses.27,28 Based on our in-silico analysis, hsa-miR-125b-5p is particularly associated with the Hippo signaling pathway and is known to modulate the expression of inflammatory cytokines, influence the inflammatory state of macrophages, and affect macrophage polarization.29,30
Lysine as well as other amino acid is also crucial for immune system function. Studies have shown that the degradation of lysine and other amino acids provides the necessary energy for immune system activation and is involved in macrophage activation, polarization, and function.31 The product of lysine degradation, pipecolic acid, is also known to regulate immune function.32 Among the miRNAs identified, hsa-miR-21-5p has the most significant effect on lysine degradation. miRNa-21 is known to influence macrophage polarization, with its deletion resulting in inhibited polarization to M2 macrophages in chronic obstructive pulmonary disease settings.18 Increased expression of hsa-miR-21 has been shown to enhance macrophage phagocytosis.33
The secretome in this study was administered via intramuscular injection (IM). IM delivery allows the secretome to be deposited directly into muscle tissue, where it can be gradually released into the bloodstream, potentially providing a sustained therapeutic effect. The rich blood supply in muscle tissue facilitates the absorption of the secretome into the circulatory system, while the large surface area of the muscle makes IM administration both straightforward and practical.34 Research has demonstrated the effectiveness of IM delivery for achieving systemic effects, with studies showing that intramuscular administration of MSCs can result in widespread benefits.35 Additionally, exosomes, a key component of the secretome, have been shown to reach systemic circulation even when administered intramuscularly.36,37
The safety of the secretome was evaluated through an acute toxicity test, which is designed to assess the tolerability of a substance following a single administration and typically provides information on the LD50, or lethal dose 50, which is the dose causing death in 50% of test subjects.38 In our study, a single administration of the secretome at the highest possible intramuscular dose did not cause any changes in behavior, physical examination results, body weight, or organ weight from 24 hours up to 15 days post-administration. Additionally, this dose did not result in any mortality or other noticeable disturbances in the mice, indicating that a single intramuscular administration of the secretome is well-tolerated.
Our study demonstrates that the secretome derived from human UCMSCs exhibits immunomodulatory activity. While the exact mechanism was not explored in this study, the miRNA pathway analysis suggests potential mechanisms. Although we observed very low expression of IL-10, further investigation of other soluble proteins may reveal additional mechanisms underlying immunomodulation. Additionally, our study confirms the safety of a single administration of the secretome. However, a limitation of our study is that the carbon clearance assay was conducted exclusively on male mice to avoid physiological variations associated with the estrus cycle in female mice.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)