Keywords
BMP15; BMPR1B; GDF9; Goat; KISS1; Litter size
This article is included in the Genomics and Genetics gateway.
BMP15; BMPR1B; GDF9; Goat; KISS1; Litter size
The prolific trait in goats refers to the ability to have multiple kids at the same time. Genetic variations that increase the rate of ovulation and the number of offspring per birth affect the prolific trait of each goat differently.1 Indonesia has a goat population of nearly 19 million, producing 72.553 tons of meat per year from 7,144,010 heads (FAOSTAT, 2019). According to the Indonesian Ministry of Agriculture (2020), goat meat accounts for 1.8% of total national meat production. The import of goat meat products reached 1.49%, with a total of 387,663 heads imported. This condition demonstrated that increasing the goat population is critical to meeting national needs.2 The genetics of prolificacy emphasizes the importance of three major fecundity genes in sheep, bone morphogenetic protein 15 (BMP15), Bone morphogenetic protein receptor type IB (BMPR1B) and growth differentiation factor 9 (GDF9). All three genes are members of the TGF superfamily.3 First identified as a fecundity gene in sheep was the booroola gene (BMPR1B). There was no change in the expression level of this gene, but the ovulation rate increased due to the cumulative impact of this substitution mutation.4 The FecB locus is autosomal with codominant expression and is characterized by “precocious” ovarian follicle differentiation, resulting in the production of a large number of ovulatory follicles that are smaller in diameter than wild-type follicles. In sheep, mouse, and rat granulosa cells, BMPR1B has been identified as one of the type 1 receptors downstream of BMP15.5
Bone morphogenetic protein 15 (BMP15), also known as FecX (fecundity chromosome X), is a gene that regulates the prolific properties of different sheep types1 paracrine factor that promotes follicle growth, granulosa cell proliferation, and cell-survival signaling, and an X-linked gene (FecX locus) of sheep belonging to the TGF family,6 BMP15 plays a role in the booroola phenotype and is additionally usually known as the GDF-9B genetic code for protein synthesis in oocytes, which further improves follicle formation and fecundity in sheep and goats. It is unknown what role BMP15 genes play in granulose cell management.7 The GDF9 gene encodes a transforming growth factor that oocytes secrete during folliculogenesis. GDF9 regulates female sexual reproduction, gonad development, gamete generation, and the ovulation cycle by modulating the signaling pathways of the transforming growth factor-β receptor and transmembrane receptor protein serine/threonine kinase.8 So far, research on the kisspeptin (KISS1) gene as a candidate gene for reproductive characteristics in animals has demonstrated that this gene plays a vital function in animal reproduction.9 Kisspeptins directly promoted the release of gonadotropin-releasing hormone (GnRH) via KiSS1R, which in turn stimulates the production of leutenizing hormone (LH) and follicle stimulating hormone (FSH). Additionally, research has shown that kisspeptin plays a part in the formation of the placenta.10 Because KISS1 is important as a regulator of puberty initiation, variations in this gene are anticipated to be associated with reproductive features in goats such as high prolificacy, sexual precocity, and year-round estrus phenotypes. Kisspeptins may have a role in photoperiodic reproductive regulation in hamsters.9 SNPs in the goat KISS1 gene were consequently found to be significantly associated with litter size.11
The current review discusses the outcomes provided in numerous research and offers data by pooling these studies' research findings. To the best of knowledge, no review studies have been carried out on the relationship of candidate genes with litter size in Indonesian goats. The purpose of this review paper was to investigate the influence of genetic variants on litter size in Indonesian goats, specifically focusing on the four genes that have received the most attention in the scientific community: BMPR1B, BMP15, GDF9, and KISS1. The review paper was constructed by combining the results of all research that had been previously published in scientific papers (Table 1).
A detailed and comprehensive search was conducted to find related research and data published between 2011 and September 2022 using critical databases including ResearchGate, Google Scholar, PubMed, Google search engine and Science direct. One reviewer (M.A) independently searched for relevant studies using the following search terms: “BMPR1B”, “BMP15”, “GDF9”, “KISS1” “goat”, “litter size”, “prolificacy”, “Indonesian goat litter size” “SNP” “polymorphism” and “fecundity”. Furthermore, the reference lists of the papers that were found were examined to identify qualified studies that might not have been found through the journal and database search processes.
Only papers reporting associations between candidate genes and litter size, sample sizes per genotype, Chi-squared means for each genotype, genotype and allele frequencies, and descriptions of statistical methods used to collect these results were included in this analysis. When there were any exclusion criteria, they were as follows: articles published in languages other than English, Conference papers, short communication papers and papers not related to animals.
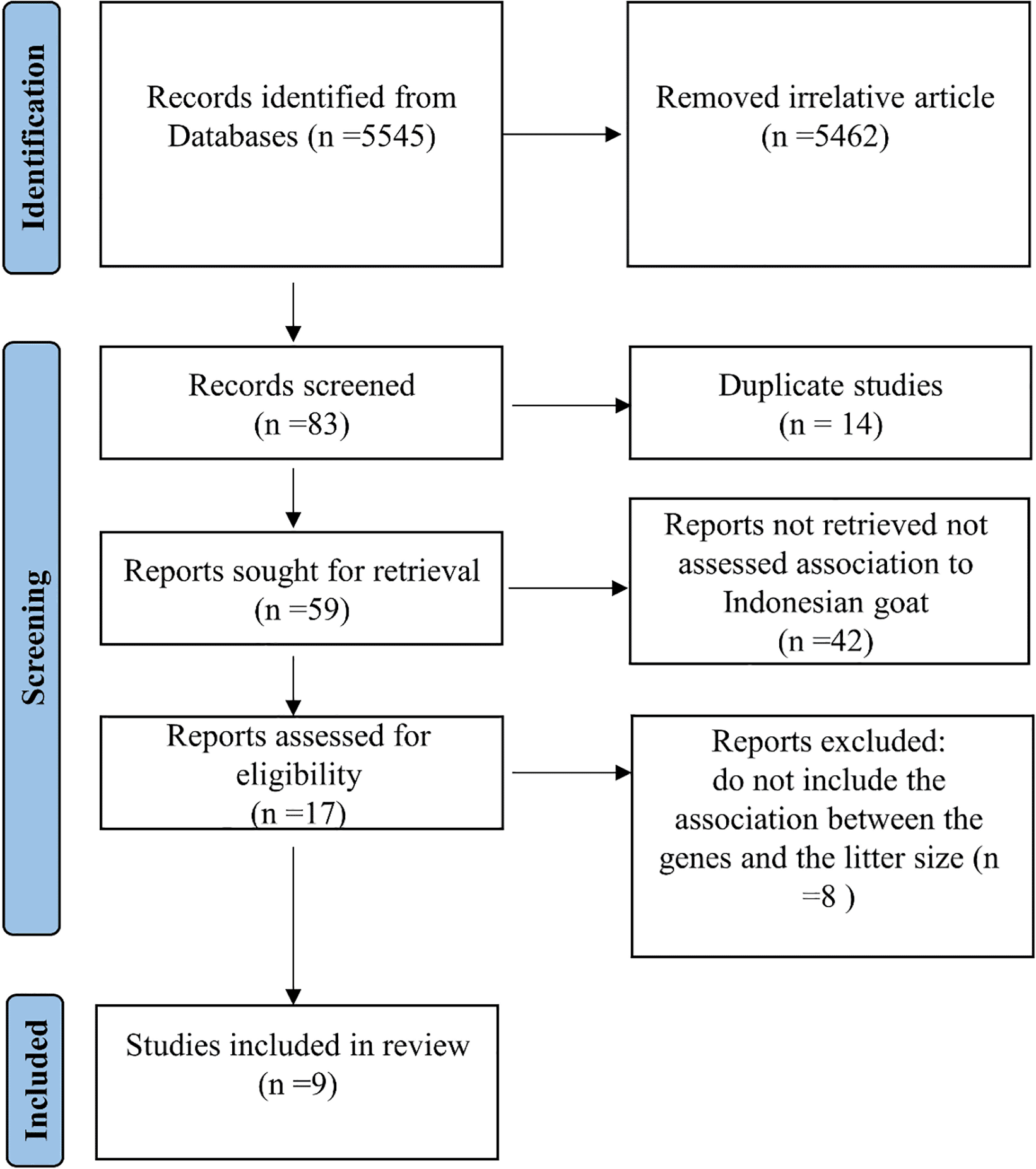
Eventually, 5545 titles of publications and papers were analyzed, with 83 being chosen for additional review. Following the removal of duplicates, the abstracts of 59 articles were chosen for screening. After reviewing the abstracts of 42 publications, the remaining 17 investigations were chosen for full paper evaluation. A further eight studies were removed after a comprehensive evaluation of the publications because they did not match our inclusion criteria. The study incorporated the remaining nine publications to guarantee high-quality data (Figure 1).
DNA polymorphism analysis can be used for a variety of purposes, including sex determination, individual paternity testing, species identification, disease detection, phylogenetic analysis, and marker-assisted selection. In comparison with conventional breeding practices, the understanding and use of DNA polymorphisms has led to the identification of unique genetic markers that may be used to precisely select animals for improved output.12 The selection of high-potential parents is critical for improving the next generation by using genetic markers associated with litter size.13 Nevertheless, due to the low selection accuracy, improving this feature is difficult. This is related to a combination of low-to-moderate heredity and the predominant effect of the environment. The utilization of molecular genetics to identify gene loci and chromosomal areas containing single nucleotide polymorphisms (SNPs) impacting commercially relevant livestock characteristics has increased at an unparalleled rate over the last decade. Marker-assisted selection technology enables the precise selection of favourable sequence variants that have been shown to improve performance, eliminating much of the guesswork associated with traditional phenotypic selection.14
Goats are known to have a number of genes that influence reproductive traits and the characteristics of their offspring, including litter size. Among these are the BMP15, KISS1, and BMPR1B genes, as well as the GDF9 gene, and all four of these genes are potential candidates for genetic marker screening of goat litter size. Through the use of genetic marker screening, a number of different polymorphisms within these genes have been identified as being associated to the performance of the reproductive system and the litter size. As a consequence of this, these SNPs have the potential to serve as novel molecular markers for determining the litter size of goat (Table 1).12–22
The SNPs of GDF9 genes were identified, and litter size traits were analyzed in four Indonesian goats (Saanen, Bligon Kacang and Kejebong). The results of the PCR restriction fragment length polymorphism (RFLP) analysis revealed four new polymorphisms in all three GDF9 genes. PCR-RFLP analysis revealed the SNP V397I with accession number XM_013965446.2,15 and three SNPs in (g.3615T>C g.3760T>C g.3855A>C) with accession numbers EF446168, EU883989 and KY780296.20 In the 60 Saanen goats tested, SNP V397I revealed only one genotype GA, showing that this SNP is monomorphic. One of the two most well-known SNPs in the GDF9 gene is G1189A, also known as p.Val397Ile/V397I. This SNP is related with varying levels of prolificacy in goats all over the world.23 To this day, the V397I SNP has been the subject of a significant amount of research in a variety of goat breeds.24 Similar to the results presented in this study, a previous investigation found that the V397I SNP was nonpolymorphic in Black Bengal Indian goats, and that study genotyped the V397I SNP.15 In contrast to the results of this investigation, the V397I SNP was discovered to exhibit polymorphism in a number of different breeds. Because the frequency of the G1189A mutant allele was 0.06 in Jamunapari goats and 0.15 in crossbred goats, this indicates that the mutation is stable in the population.24
Only the SNP g.3855A>C was analyzed in the second investigation to determine genotype frequencies, which discovered only two genotypes AA and AC, with the genotype CC not detected in these three goats (Bligon, Kacang, and Kejebong). Because the SNPs found in goats in those studies exhibited low polymorphism, no further research was undertaken to investigate whether it relates to litter size in goats.
There has been an evaluation of polymorphism of the BMP15 gene in the litter size and there was the detection of SNPs associated with litter size traits of c.38A>G and c.49G>A,19 746A>G,1 g.735A>G16 and g.135G>C17 with accession numbers EU743938.1, JQ350891.1, JQ320890 and AF236078 respectively, out of Boer, Kacang, Boerka, Kosta, Samsoir an Etawah breed. These SNPs of the BMP15 gene were identified using DNA pool sequencing and PCR-RFLP methods. The findings of the BMP15 gene sequence with GenBank (access code EU74393.1) revealed that the genotype frequency of the BMP15 gene based on Boer goat, Kacang of samples was found by two SNP, c.38A>G and c.49G>A.
The outcome of the genotype frequency on SNP c.49G>A, the genotype frequency of the BMP15 gene based on samples of Boer, Kacang, and Boerka goats used in this study presented the following genotypes: GG, GA, and AA (Table 2).19 Boer goat genotypes found in BMP15 SNP c.49G>A genes were GG, GA, and AA, with genotype frequencies of 0.471, 0.235, and 0.294. Kacang goat genotypes reported in BMP15 SNP c.49G>A genes have been GG, GA, and AA, with genotype frequencies of 0.125, 0.438, and 0.438, respectively. Boer goat genotypes found in the BMP15 SNP c.49G>A gene were GG, GA, and AA, with genotype frequencies of 0.235, 0.235, and 0.529, respectively. Additionally, the heterozygosis (Ho) and heterozygosity (He) tests demonstrated that the observed Ho in Boer, Kacang, and Boerka goats was not significantly different from the expected He. The difference between the observed heterozygosis value and heterozygosis may be utilized to detect a genetic inequality in the observed Boer goat, Kacang, and Boerka populations, showing that selection activity has already happened, and that random marriage has not occurred. The detected heterozygosis value (Ho) of the BMP15 gene in Boer, Kacang and Boerka goats was 0.235, 0.438, and 0.235, respectively. On SNP c.49G>A, the greatest heterozygosis value in Kacang goats is 0.438. The population balance is presented in Table 2 by the Hardy–Weinberg equilibrium (HWE). The results demonstrate that the BMP15 gene on SNP c.49G>A is in a balanced state.
| Author | Goat number | Genotype frequencies | Allele frequencies | He | χ2 | Littersize (mean±SE) | P value | |||
|---|---|---|---|---|---|---|---|---|---|---|
| 19 | Boer (17) Kacang (16) Boerka (17) | GG 0.471 0.125 0.235 | GA 0.235 0.438 0.235 | AA 0.294 0.438 0.529 | G 0.588 0.344 0.353 | A 0.412 0.656 0.647 | 0.484 0.451 0.457 | 4.496 0.015 3.996 | - | - |
| 1 | Local goat (100) | GG 0.00 | G+ 0.07 | ++ 0.93 | G 0.035 | + 0.965 | 0.070 | 0.132 | 1.65±0.2a | P<0.05 |
| 2 | Kacang (30) Kejobong (30) Senduro (30) | CC 0.71 0.38 1.00 | CG 0.14 0.12 0.00 | GG 0.14 0.50 0.00 | C 0.79 0.44 1.00 | G 0.21 0.56 0.00 | 0.33 0.22 0.00 | 2.32 4.5D 8.0D | 2.67±0.13a 2.33±0.25 N/A | P<0.004 |
| 2 | Kacang (30) Kejobong (30) Senduro (30) | GG 0.29 0.75 0.38 | GA 0.43 0.12 0.62 | AA 0.29 0.13 0.00 | G 0.50 0.81 0.67 | A 0.50 0.19 0.33 | 0.5 0.3 0.43 | 0.14 2.78 1.65 | 3.00±0.0a 3.00±0.0a 3.00±0.0a | P<0.002 |
| 22 | Kaligesing (48) | TA 0.6 | AA 0.40 | TT 0.00 | A 0.7 | T 0.3 | - | - | - | - |
| 15 | Saanen (60) | GA 1.00 | AA 0.00 | GG 0.00 | G 1.00 | A 0.00 | - | - | - | - |
| 16 | Gembrong (5) Kosta (4) Samosir (8) Kacang (10) | AA 0.40 1.00 0.88 0.60 | GG 0.2 0.00 0.00 0.00 | AG 0.4 0.00 0.12 0.4 | A 0.60 1.00 0.94 0.80 | G 0.40 0.00 0.06 0.20 | - | 0.14 0.00 0.04 0.62 | - | - |
| 17 | Etawah (106) | CC 0.00 | CG 0.00 | GG 1.00 | C 0.00 | G 1.00 | - | - | - | - |
| 17 | Etawah (106) | GG 0.038 | CG 0.00 | CC 0.962 | G 0.962 | C 0.038 | - | - | 1.75±0.96a 1.62±0.58a | P>0.05 |
| 17 | Etawah (106) | AA 0.019 | AT 0.179 | TT 0.802 | A 0.11 | T 0.89 | - | - | 1.59±0.58a 1.79±0.63a 1.50±0.71a | P>0.05 |
| 18 | Kacang (7) Kejobong (8) Senduro (8) | AA N/A | AG N/A | GG N/A | A N/A | G N/A | - | - | 4.01±0.96a | 0.0027 |
| 20 | Bligon (10) Kacang (2) Kejobong (2) | CC 0.00 | AC 0.071 | AA 0.929 | A 0.96 | C 0.04 | - | - | - | - |
The analysis of the diversity of the BMP15 SNP 746A>G gene1 produced + and G alleles which had ++ and G+ genotypes, table 2 shows that the GG genotype was not found in local goats. Table 2 also shows that the frequency of the + allele in the BMP15 gene is higher compared with the G allele. The + allele frequency is 0.965 and the G allele frequency is 0.035, indicating that the + allele frequency is greater than the G allele frequency in the BMP15 gene. This indicates that the candidate gene BMP15, which was discovered in various types of sheep, was also discovered in local goats, with a value of more than 1%. Thus, the gene BMP15 in local goats is polymorphic because the allele frequency value is greater than 1%. The Chi-squared (χ2) test analysis against the BMP15 gene revealed that the genotype and allele frequencies are homogeneous. These findings show that the local goat population is still healthy and balanced HWE, and that the goat population has never been selected. The observed heterozygosity value (He) in Table 2 is 0.070 greater than the expected heterozygosity value (He) of 0.068. This finding indicates that the observed and expected heterozygosity values in local goats do not differ significantly in the BMP15 gene fragment.
The result of SNP 735A>G16 (Table 2) reveals allele and genotype frequencies. Gembrong goats come in all three genotypes. The GG genotype, on the other hand, was missing in Kosta, Samosir, and Kacang goats. This result showed that allele A is higher than allele G in all goats. The Kosta goat had only homozygous AA, the Samosir goat had two genotypes AA (0.88) and AG (0.12), and three genotypes were found in Gembrong as the following AA (0.40), GG (0.20), and AG (0.40). Meanwhile, in the Kacang goat two genotypes were detected in the study24 AA and AG were 0.58 for A allele and G allele was 0.42, in contrary Das et al.25 reported that allele G had higher frequency than allele A as the following 0.762 and 0.238 respectively. PCR-RFLP analysis in SNP (g. 135G>C)17 showed only the GG genotype. The DNA observed was monomorphic for the BMP15 gene (exon 1) with GG genotype (Table 2).
There has been an evaluation of polymorphism of the BMPR1B gene in the litter size and there was the detection of SNPs associated with litter size traits of c.85A<G, c.742C<T, c.743T<C, c.744C<T and c.225 G<C with accession number KC142198.1.17 the study samples were collected from the Etawah breed. These SNPs of the BMPR1B gene were identified using PCR-RFLP methods. The frequency of the C allele in the gene BMPR1B (exon 1), was found to be higher than G (Table 2). The genotype frequencies found in the BMPR1B SNP c.225 G<C gene were GG and CC with genotype frequencies of 0.038 and 0.962. Meanwhile, allele frequencies were 0.038 and 0.962 for G and C, respectively. In this study, a heterozygous genotype was not found, while both C and G genes were not fixed. The condition of the BMPR1B gene (exon 1) in research causes the Hardy–Weinberg balance is not met (P<0.05). By contrast, after researching the FecB mutation in six distinct breeds of productive goats (Black Bengal, Beetal, Barbari, Malabari, and Sikkim), the researchers found that none of the goats were homozygous carriers of the trait.26
The polymorphism of the KISS1 gene in the litter size was evaluated, and it was discovered of 14 SNPs in Kacang, Kejobong and Senduro as the following (g.2064T>A, g.250G>A, Ng.2540C>T, g.2196G>C, g.2270C>T, g.2425C>G, g.2436A>G, g.2360A>G, g.2459G>A, g.1978C>T, g.2055A>C, g.2489T>C, g.2601T>A),2 For the polymorphism research analysis, only two SNPs, g.2425C>G and g.2459G>A, were explored utilizing PCR sequencing from three population groups in Kejobong Senduro and Kacang. Polymorphisms could not be detected in Senduro goats, whilst Kacang and Kejobong were polymorphic, with genotype frequencies in Kacang CC 0.71, CG0.14, and GG 0.14, and allele frequencies of C 0.79, G 0.21, and 0.50, respectively, with the allele frequencies for allele C, were 0.44 and 0.56, respectively. In the case of SNP g.2459G>A, polymorphism was found in all breeds, with genotypes GG (0.29, 0.75, and 0.38), and GA (0.43, 0.12, and 0.62) in the three breeds, while genotype AA was only found in two breeds (Kacang and Kejobong), with genotypes AA (0.43, 0.12, and 0.62). (0.29 and 0.13). In the Damascene and Zarabi goat breed, those with the TT genotype at intron 1 had a greater progesterone level compared to those with the TA genotype.9 In the meanwhile, it was discovered that there was an SNP known as T125A in the intron 1 region of the KISS1 gene. The allelic frequency of alleles A and B was measured at 0.43 and 0.57, respectively.27
In both SNPs, Kacang goats were in HWE. Kejobong and Senduro goats were not subjected to HWE at g.2425C>G, but rather at g.2459G>A (χ2>3.841). SNP T125A was identified in 48 Kaligesing goats aged 3 years old using PCR-RFLP and DNA sequencing, and the results revealed that The KISS1 gene was polymorphic in the studied population, having one single nucleotide variation. (SNP T125A). SNP was then used to study the genotype of all individuals using the PCR-RFLP method. There were three genotypes identified (TT, TA, and AA). The genotype frequency of TA was 0.60, AA was 0.40, and TT was not included in the genetic diversity calculation. T and A allele frequencies were 0.30 and 0.70, respectively. The SNP genotype distribution differed from the HWE (χ2=8.10; P=0.025).
Mulyono et al.17 had investigated KISS1 gene in goats using PCR-RFLP, four SNPs were found in exon 1 and intron 1 of the KISS1 gene (c.585A>C, c.50G>A, c.82A>T, c.114C>A, c.116C>A, and c.57T>C). In this review study, only the genotype and allele frequency of the SNP in intron 1 was examined. In the Eatawah goat, there have been three genotypes (AA, AT, and TT), the genotype frequencies for the A and T alleles were 0.11 and 0.89, respectively, whereas the genotype frequencies for the AT and TT genotypes were 0.179 and 0.802.16 Indicated that if a polymorphic locus has 0.05, it is classified as an SNP. The allele and genotype frequencies will maintain unchanged as long as there will be no selection, mutation, non-random mating, migration, or genetic drift in the population.
In Table 2, a summary of the litter sizes of a few breeds of Indonesian goats was provided. In the study conducted by Mulyono et al.,17 it was revealed that combinations of alleles in the BMP15 gene produced two genotypes: genotype ++ with a frequency of 0.93 and the average of litter size of 1.59±0.58, and genotype G + with a frequency of 0.07 and an average litter size of 1.65±0.2 similar outcomes shown in SNPs of 818C>T, and 959A>C.5 However, research investigating how BMP15 mutations affect litter size is limited. Used PCR-SSCP and DNA sequencing to analyze exon 2 of the BMP15 gene in two Chinese local goat breeds. According to the findings of the research, Funiu white goats had three genotypes (AA, BB, and AB), whereas Taihang black goats only had two genotypes (AB and BB). Those Funiu white goats carrying the genotype BB had either 0.91 or 0.82 more kids than those carrying the AB or AA genotypes, correspondingly, indicated that the BMP15 gene would be a significant contributor to the fertility of Funiu white goats.28
The result of the KISS1 gene with SNP (g.2459C>G) showed that the litter size of the same genotype varied between the two studies with different flocks of Kacang, Kejobong and Senduro2,18 The first study showed the same result of litter size 3.00±0.0 among the three breeds, whereas the second study found a larger litter size (4.01±0.96) in KISS1 (g.2459C>G) with the same breeds. This could be due to the small number of samples used in the second study, but it was also discovered that the associated SNP g.2425C>G had a significant effect on litter size in the KISS1 gene among the Kacang and Kejobong breeds, with values of 2.67±0.13 and 2.33±0.25, respectively. Febriana2 found four unique SNPs in the KISS1 gene intron 1, of which two are prominently associated with reproductive traits in local goat breeds native to Indonesia. Both the AA genotype at g.2436A>G and the GA genotype at g.2436A>G had a significant influence on litter size across all Indonesian native goat breeds. On the other hand, the genotypes g.2360A>G, g.2425C>G, and g.2510G>A did not exhibit the same link with LS in various breeds of goats. It's interesting to note that the Kacang and Kejobong breeds are the only ones where the AG genotype at g.2360A>G, CC genotype at g.2425C>G, and AA genotype at g.2510G>A showed greater LS. In addition, the CC genotype at the g.2425C>G locus in SD goats has nearly the same LS average as Kacang (2.67±0.23) and is higher than Kejobong (2.33±0.25). However, g.2425C>G and g.2459G>A both expressed that the genotype at these loci has a substantial impact on the LS at the first and the third parities, in contrast to the second parity. Nonetheless, the GA genotype at g.2459G>A was only significant in the first parity. In addition, the CC genotype at g.2425C>G and the AA genotype at g.2436A>G revealed a relationship with LS at the third parity. This association was substantially higher than the association with LS at the second parity and the first parity. At the third and last parity, we were, however, lacking in samples. In general, the CC genotype at locus g.2425C>G, the AA genotype at locus g.2436A>G, and the GA genotype at locus g.2459G>A were good genotypes related with LS and parity substantially (p>0.05) in local goat breeds from Indonesia.2 In contrast, there was no interaction impact between parity and genotype. on litter size in Black Bengal goats.29 On the other side, in the fourth parity, the TT genotype was connected to greater litter size in Chinese goat breeds than the CC genotype was. This was revealed by evaluating the litter size values between the two genotypes.11 It was discovered that there is a substantial correlation between the genotypes of Gaddi goats and the number of kids born to each litter. The results of the research indicate, on the other hand, that there is a connection between the allele T found in the KISS1 gene and the size of the litter.27 The polymorphism in the goat KISS1 gene was found to have a substantial influence on the reproductive features in a prior study.9 Additionally, in Guangzhong, Saneen, and Xinong goats, a discernible link was found to exist between the KISS1 gene polymorphism and the litter size,11 a further SNP in the KISS1 gene was discovered in the Guanzhong breed of goat that was associated with litter size.30–32 Research conducted on Egyptian goat breeds (Zaraibi, Baladi, and Barki) indicated that two SNPs in the KISS1 promoter region were substantially linked with litter size. These SNPs were T2124A and C2270T. According to the findings of 29, there are three polymorphisms in Black Bengal goats that are related to larger litter sizes. These are G296C, G2510A, and C2540T. In the promoter region of the KISS1 gene, a new polymorphism called G231C was discovered. This polymorphism was found to have a statistically significant connection with the litter size in goats (P<0.05).10
Study of Exon 1 in gene BMPR1B with SNP c.225 G>C showed that genotype GG had a higher litter size than the other genotypes, the study had found no association among genotypes of BMPR1B (exon 1) gene and litter size.17 Relevant to the results that obtained, the surveyed BMPR1B mutation did not have a major links with litter size in Markhoz goats. Additionally, goat breeds from India, Iran, China, Egypt, and Malaysia were examined for the FecB mutation, and all of these goats were determined to be homozygous non-carriers of the mutation.5 There is no evidence to suggest that the BMPR1B gene is linked to any reproductive traits in goats,33 in a sample of nine Indian goat breeds, researchers discovered two unique SNPs in the promoter region of the BMPR1B gene, 242T>C and 623G>A, but also no association with prolificacy traits.34
One of the few studies to identify a correlation between GDF9/V397I and litter size in goats35 indicated that V397I has a substantial influence on litter size; furthermore, it found related findings in Chinese dairy goats.36 Contrastingly, neither Q320P nor V397I were associated with litter size in seven native Indian goat breeds investigated.35 Wang et al.23 found that V397I had a significant impact on litter size until sample size exceeds 1300. We didn't find any places in the GDF9 gene that were strongly linked to the size of a Black Bengal goat's litter. According to Ahlawat et al.,3 the impact of parity on litter size in Indian Black Bengal goats is significant (P<0.01). Das et al.37 also noticed a highly significant (P<0.001) influence of parity on litter size in this investigation. Ahlawat et al.5 investigated the relationship with new SNPs in the BMP15, BMPR1B and GDF9 genes and litter size in seven Indian goat breeds. No SNPs including 1189G>A and 959A>C in GDF9 and 242T>C in BMPR1B were found to be linked with litter size. According to the findings in Indonesian goats with the GDF9/V397I and all over the world, it was concluded that there is not likely to be a substantial association between the SNPs in goats as there is in sheep. Additional research with larger samples may provide additional light on the connection between this SNP and goats.
The discovery of polymorphisms in fecundity genes has aided in the selection of goats with superior reproductive performance, as a result of which goat litter size has increased. In this review, 36 SNP loci from the goat BMP15, KISS1, BMPR1B, and GDF9 genes were gathered and sorted, the relationship between part-potential SNPs and the trait of goat litter size was analyzed and discussed. Among these mutations, four were monomorphic: GDF9/V397I in Saanen, KISS1/g.2425C>G in Senduro, BMP15/g.135G>C in Etawah, and BMP15/g.735A>G in Kosta. Meanwhile, these six SNPs were studied for their association with litter size traits in the Indonesian goat breeds (Table 2). In Indonesian goats, only four of the six SNPs were shown to be substantially associated with litter size traits (Table 2). The goats used in this study were reared or raised on institutions' farms and were chosen for their high performance of desirable traits such as productivity. As a result, there is a good chance that the SNPs were already selected for the genotype associated with high reproduction performance expression. As a result, additional research should be conducted with larger sample size and with different small and large farms from different regions and areas.
Overall, the findings in the Indonesian goat support the notion that the BMP15, KISS1, BMPR1B, and GDF9 genes increase litter size. These outcomes are critical for developing molecular breeding strategies for goats as well as improving goat reproduction and production performance.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)