Keywords
Uniprot ID P31483, TIA1, RNA-binding protein TIA1, antibody characterization, antibody validation, Western Blot, immunoprecipitation, immunofluorescence
This article is included in the YCharOS (Antibody Characterization through Open Science) gateway.
This article is included in the Cell & Molecular Biology gateway.
Uniprot ID P31483, TIA1, RNA-binding protein TIA1, antibody characterization, antibody validation, Western Blot, immunoprecipitation, immunofluorescence
In this new version of the article, additional references have been included in the introduction to provide more extensive background on the role of TIA1 in various diseases. Additionally, in the Results & Discussion section, we clarify the YCharOS initiative as a public resource for the scientific community and encourage readers to use the data provided as a guide to selecting high-performing antibodies for their specific needs.
See the authors' detailed response to the review by Jozsef Gal
T-cell intracellular antigen 1, or TIA1, is a cytotoxic granule-associated RNA-binding protein involved in regulating alternative pre-mRNA splicing and mRNA translation when bound to 3’ uridine-rich RNA sequences.1–4 Suppressing translation in environmentally stressed cells and promoting stress granule formation, TIA1 modulates cellular response to stress and inflammation.5,6
Comparable to other RNA-binding proteins, disrupting the function of TIA1 can lead to various diseases including cancer,7–9 autoimmune diseases10 and neurodegenerative disorders.11–14 Studies have demonstrated that mutations to the TIA1 gene may delay the disassembly of stress granule, resulting in insoluble and immobile stress granules, a clinical feature of ALS and FTD.6,11 Significant efforts are required to further elucidate the relationship between dysregulated RNA metabolism and ALS/FTD pathogenesis which may lead to novel therapeutic discoveries.11,15
Mechanistic studies would be greatly facilitated with the availability of high-quality antibodies. Here, we compared the performance of a range of commercially-available antibodies for TIA1 and identified high-performing antibodies for Western Blot, immunoprecipitation and immunofluorescence, enabling biochemical and cellular assessment of TIA1 properties and function.
Our standard protocol involves comparing readouts from wild-type (WT) and knockout (KO) cells.16–23 The first step was to identify a cell line(s) that expresses sufficient levels of TIA1 to generate a measurable signal. To this end, we examined the DepMap transcriptomics database to identify all cell lines that express the target at levels greater than 2.5 log2 (transcripts per million “TPM” + 1), which we have found to be a suitable cut-off (Cancer Dependency Map Portal, RRID:SCR_017655). Commercially available HAP1 cells expressed the TIA1 transcript at RNA levels above the average range of cancer cells analyzed. Parental and TIA1 KO HAP1 cells were obtained from Horizon Discovery (Table 1).
| Institution | Catalog number | RRID (Cellosaurus) | Cell line | Genotype |
|---|---|---|---|---|
| Horizon Discovery | C631 | CVCL_Y019 | HAP1 | WT |
| Horizon Discovery | HZGHC003048C010 | CVCL_TS30 | HAP1 | TIA1 KO |
For Western Blot experiments, we resolved proteins from WT and TIA1 KO cell extracts and probed them side-by-side with all antibodies in parallel (Figure 1).17–23
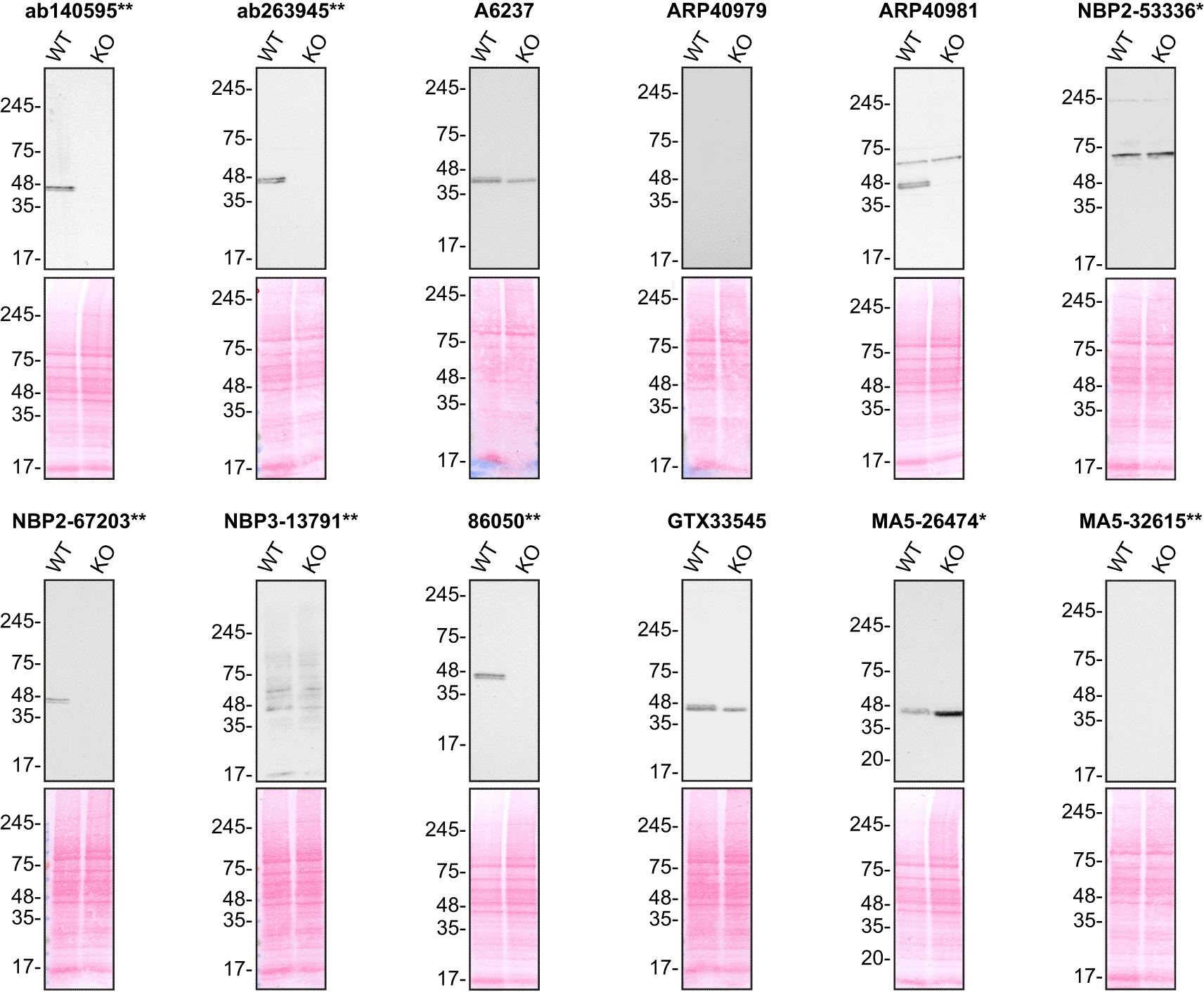
Lysates of HAP1 (WT and TIA1 KO) were prepared and 50 μg of protein were processed for Western Blot with the indicated TIA1 antibodies. The Ponceau stained transfers of each blot are presented to show equal loading of WT and KO lysates and protein transfer efficiency from the acrylamide gels to the nitrocellulose membrane. Antibody dilutions were chosen according to the recommendations of the antibody supplier. Antibody dilution used: ab140595** at 1/5000; ab263945** at 1/1000; A6237 at 1/1000; ARP40979 at 1/500; ARP40981 at 1/200; NBP2-53336* at 1/1000; NBP2-67203** at 1/1000; NBP3-13791** at 1/500; 86050** at 1/1000; GTX33545 at 1/500; MA5-26474* at 1/2000; and MA5-32615** at 1/500. Predicted band size: 43 kDa. *=monoclonal antibody, **=recombinant antibody.
For immunoprecipitation experiments, we used the antibodies to immunopurify TIA1 from HAP1 cell extracts. The performance of each antibody was evaluated by detecting the TIA1 protein in extracts, in the immunodepleted extracts and in the immunoprecipitates (Figure 2).17–23
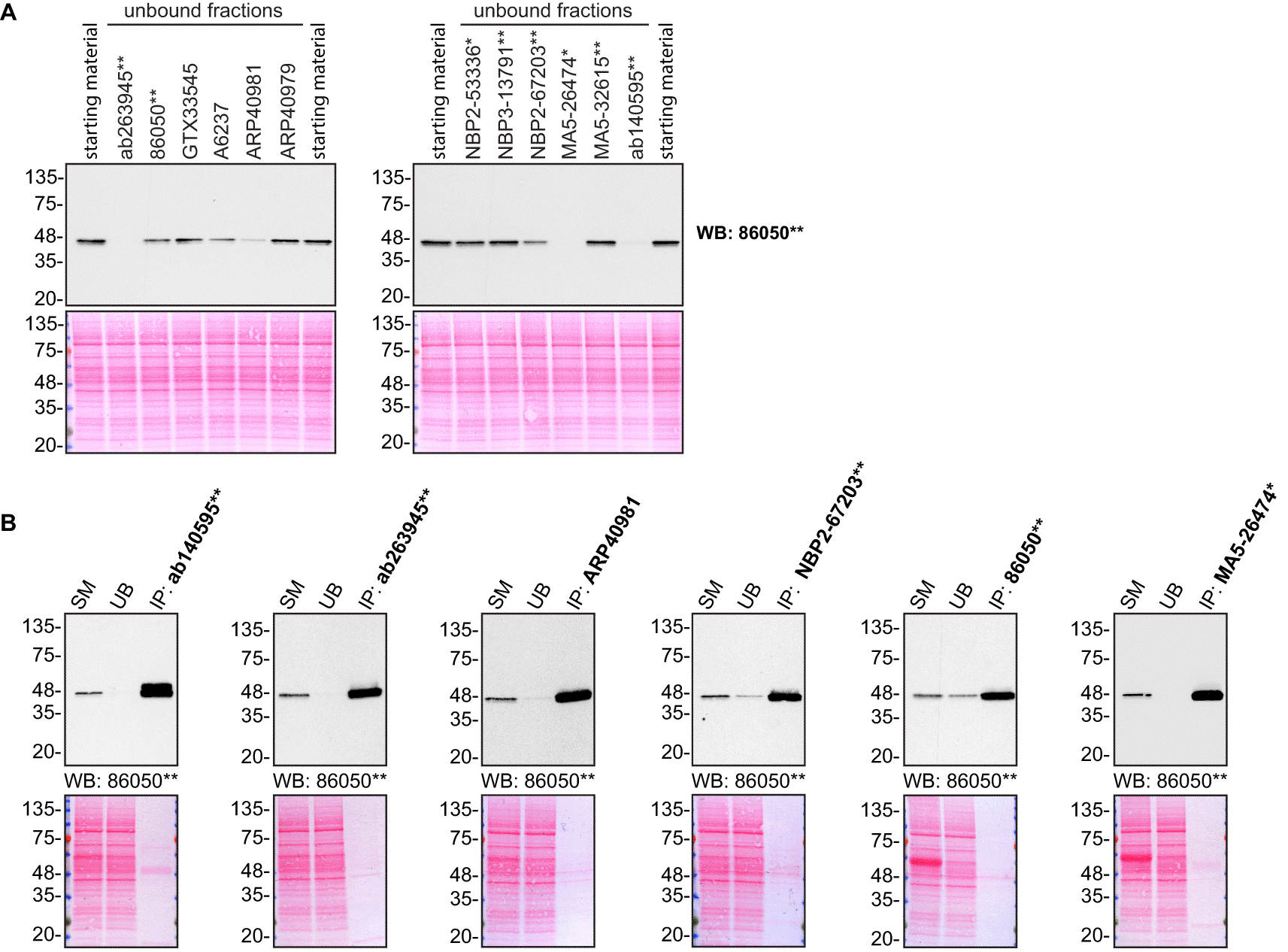
HAP1 lysates were prepared, and IP was performed using 1.0 μg of the indicated TIA1 antibodies pre-coupled to Dynabeads protein G or protein A. (A) Ability of the antibodies to capture TIA1 was assessed by comparing the level of protein available in the starting material to the level remaining in the unbound fraction. (B) The immunoprecipitates for antibodies which could immunocapture TIA1 in (A) are shown. For Western Blot, 86050** was used at 1/1000 in A) and B). The Ponceau stained transfers of each blot are shown. SM=4% starting material; UB=4% unbound fraction; IP=immunoprecipitate. *=monoclonal antibody, **=recombinant antibody.
For immunofluorescence, as described previously, antibodies were screened using a mosaic strategy.24 In brief, we plated WT and KO cells together in the same well and imaged both cell types in the same field of view to reduce staining, imaging and image analysis bias (Figure 3).
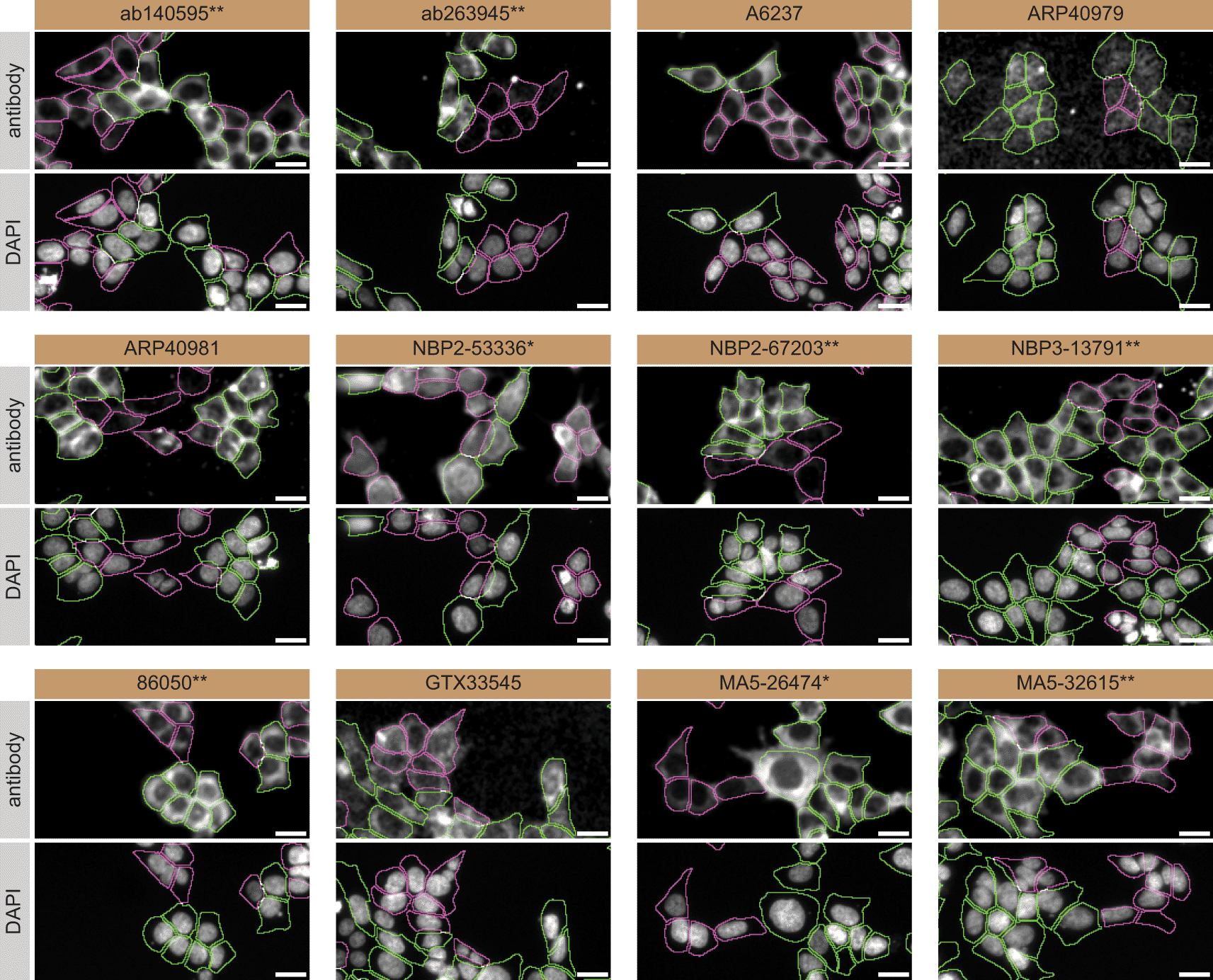
HAP1 WT and TIA1 KO cells were labelled with a green or a far-red fluorescent dye, respectively. WT and KO cells were mixed and plated to a 1:1 ratio in a 96-well plate with optically clear flat-bottom. Cells were stained with the indicated TIA1 antibodies and with the corresponding Alexa-fluor 555 coupled secondary antibody including DAPI. Acquisition of the blue (nucleus-DAPI), green (identification of WT cells), red (antibody staining) and far-red (identification of KO cells) channels was performed. Representative images of the merged blue and red (grayscale) channels are shown. WT and KO cells are outlined with green and magenta dashed line, respectively. Antibody dilutions were chosen according to the recommendations of the antibody supplier. Exceptions were given to antibodies ab140595**, A6237, NBP2-67203**, and GTX33545, which were titrated to their respective concentrations found bellow, as the signals were too weak when following the suppliers' recommendations. When the concentrations were not indicated by the supplier, which was the case for antibodies ab263945**, ARP40979, ARP40981, and NBP3-13791** we tested antibodies at 1/300, 1/500, 1/500 and 1/200, respectively. At these concentrations, the signal from each antibody was in the range of detection of the microscope used. Antibody dilution used: ab140595** at 1/600; ab263945** at 1/300; A6237 at 1/1000; ARP40979 at 1/500; ARP40981 at 1/500; NBP2-53336* at 1/100; NBP2-67203** at 1/1000; NBP3-13791** at 1/200; 86050** at 1/100; GTX33545 at 1/800; MA5-26474* at 1/1000; and MA5-32615** at 1/100. Bars = 10 μm. *=monoclonal antibody, **=recombinant antibody.
In conclusion, we have screened TIA1 commercial antibodies by Western Blot, immunoprecipitation and immunofluorescence. Several high-quality antibodies that successfully detect TIA1 under our standardized experimental conditions can be identified. In our effort to address the antibody reliability and reproducibility challenges in scientific research, the authors recommend the antibodies that demonstrated to be underperforming under our standard procedure be removed from the commercial antibody market. However, the authors do not engage in result analysis or offer explicit antibody recommendations. A limitation of this study is the use of universal protocols - any conclusions remain relevant within the confines of the experimental setup and cell line used in this study. Our primary aim is to deliver top-tier data to the scientific community, grounded in Open Science principles. This empowers experts to interpret the characterization data independently, enabling them to make informed choices regarding the most suitable antibodies for their specific experimental needs. Guidelines on how to interpret antibody characterization data found in this study are featured on the YCharOS gateway.25 The underlying data can be found on Zenodo open access repository.26,27
All tested TIA1 antibodies are listed in Table 2, together with their corresponding Research Resource Identifiers, or RRID, to ensure the antibodies are cited properly.28 Peroxidase-conjugated goat anti-rabbit and anti-mouse antibodies are from Thermo Fisher Scientific (cat. number 65-6120 and 62-6520). Alexa-555-conjugated goat anti-rabbit and anti-mouse secondary antibodies are from Thermo Fisher Scientific (cat. number A21429 and A21424).
| Company | Catalog number | Lot number | RRID (Antibody Registry) | Clonality | Clone ID | Host | Concentration (μg/μl) | Vendors recommended applications |
|---|---|---|---|---|---|---|---|---|
| Abcam | ab140595** | GR223312-17 | AB_2687963 | recombinant-mono | EPR9304 | rabbit | 0.665 | Wb, IP, IF |
| Abcam | ab263945** | GR3297000-3 | AB_2885132 | recombinant-mono | EPR22999-80 | rabbit | 0.599 | Wb, IP |
| Abclonal | A6237 | 18980101 | AB_2766845 | polyclonal | - | rabbit | 2.920 | Wb, IF |
| Aviva Systems Biology | ARP40979 | QC10247-90408 | AB_2048416 | polyclonal | - | rabbit | 0.500 | Wb |
| Aviva Systems Biology | ARP40981 | QC10249-41152 | AB_938457 | polyclonal | - | rabbit | 0.500 | Wb, IP |
| Bio-Techne | NBP2-53336* | 7072-1p210821 | AB_2885158 | monoclonal | TIA1/1313 | mouse | 0.200 | Wb, IF |
| Bio-Techne | NBP2-67203** | H00823 | AB_29277441 | recombinant-mono | JM42-11 | rabbit | 1.000 | Wb, IP, IF |
| Bio-Techne | NBP3-13791** | 7072-2P210904 | AB_29277431 | recombinant-mono | TIA1/1352R | rabbit | 0.200 | other |
| Cell Signaling Technology | 86050** | 1 | AB_2800070 | recombinant-mono | D1Q3K | rabbit | 0.002 | Wb, IP |
| GeneTex | GTX33545 | 822104511 | AB_2887719 | polyclonal | - | rabbit | 0.820 | Wb, IF |
| Thermo Fisher Scientific | MA5-26474* | VL3152368 | AB_2725518 | monoclonal | OTI1D7 | mouse | 1.000 | Wb, IF |
| Thermo Fisher Scientific | MA5-32615** | VL3152613 | AB_2809892 | recombinant-mono | JM42-11 | rabbit | 1.000 | Wb, IP, IF |
Both HAP1 WT and TIA1 KO cell lines used are listed in Table 1, together with their corresponding RRID, to ensure the cell lines are cited properly.29 Cells were cultured in DMEM high-glucose (GE Healthcare cat. number SH30081.01) containing 10% fetal bovine serum (Wisent, cat. number 080450), 2 mM L-glutamine (Wisent cat. number 609065), 100 IU penicillin and 100 μg/mL streptomycin (Wisent cat. number 450201).
Western Blots were performed as described in our standard operating procedure.30 HAP1 WT and TIA1 KO were collected in RIPA buffer (25mM Tris-HCl pH 7.6, 150mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS) (Thermo Fisher Scientific, cat. number 89901) supplemented with 1x protease inhibitor cocktail (MilliporeSigma, cat. number 78429). Lysates were sonicated briefly and incubated for 30 min on ice. Lysates were spun at ~110,000 x g for 15 min at 4°C and equal protein aliquots of the supernatants were analyzed by SDS-PAGE and Western Blot. BLUelf prestained protein ladder from GeneDireX (cat. number PM008-0500) was used.
Western Blots were performed with precast midi 4-20% Tris-Glycine polyacrylamide gels from Thermo Fisher Scientific (cat. number WXP42012BOX) ran with Tris/Glycine/SDS buffer from Bio-Rad (cat. number 1610772), loaded in Laemmli loading sample buffer from Thermo Fisher Scientific (cat. number AAJ61337AD) and transferred onto nitrocellulose membranes. Proteins on the blots were visualized with Ponceau S staining (Thermo Fisher Scientific, cat. number BP103-10) which is scanned to show together with individual Western Blot. Blots were blocked with 5% milk for 1 hr, and antibodies were incubated overnight at 4°C with 5% bovine serum albumin (BSA) (Wisent, cat. number 800-095) in TBS with 0,1% Tween 20 (TBST) (Cell Signalling Technology, cat. number 9997). Following three washes with TBST, the peroxidase conjugated secondary antibody was incubated at a dilution of ~0.2 μg/mL in TBST with 5% milk for 1 hr at room temperature followed by three washes with TBST. Membranes were incubated with Pierce ECL from Thermo Fisher Scientific (cat. number 32106) prior to detection with the HyBlot CL autoradiography films from Denville (cat. number 1159T41).
Immunoprecipitation was performed as described in our standard operating procedure.31 Antibody-bead conjugates were prepared by adding 1 μg of antibody to 500 μL of Pierce IP Lysis Buffer from Thermo Fisher Scientific (cat. number 87788) in a 1.5 mL microcentrifuge tube, together with 30 μL of Dynabeads protein A- (for rabbit antibodies) or protein G- (for mouse antibodies) from Thermo Fisher Scientific (cat. number 10002D and 10004D, respectively). Tubes were rocked for ~1 hr at 4°C followed by two washes to remove unbound antibodies.
HAP1 WT were collected in Pierce IP buffer (25 mM Tris-HCl pH 7.4, 150 mM NaCl, 1 mM EDTA, 1% NP-40 and 5% glycerol) supplemented with 1x protease inhibitor (Millipore Sigma, cat. number P8340). Lysates were rocked 30 min at 4°C and spun at 110,000 x g for 15 min at 4°C. 0.5 mL aliquots at 2.0 mg/mL of lysate were incubated with an antibody-bead conjugate for ~1 hr at 4°C. The antibody-bead conjugate can elute the protein of interest if the tested antibodies can successfully bind their antigen. The unbound fractions were collected and removed while the beads were subsequently washed three times with 1.0 mL of or IP lysis buffer and processed for SDS-PAGE and Western Blot on a precast midi 4-20% Tris-Glycine polyacrylamide gels. Prot-A: HRP (MilliporeSigma, cat. number P8651) was used as a secondary detection system at a dilution of 0.3 μg/mL for experiments where rabbit antibodies are used for both immunoprecipitation and its corresponding Western Blot.
Immunofluorescence was performed as described in our standard operating procedure.17–24 HAP1 WT and TIA1 KO were labelled with a CellTrackerTM green (Thermo Fisher Scientific, cat. number C2925) or CellTrackerTM deep red (Thermo Fisher Scientific, cat. number C34565) fluorescence dye, respectively. WT and KO cells were plated in 96-well plate with optically clear flat-bottom (Perkin Elmer, cat. number 6055300) as a mosaic and incubated for 24 hrs in a cell culture incubator at 37oC, 5% CO2. Cells were fixed in 4% paraformaldehyde (PFA) (Beantown chemical, cat. number 140770-10ml) in phosphate buffered saline (PBS) (Wisent, cat. number 311-010-CL) for 15 min at room temperature and then washed 3 times with PBS. Cells were permeabilized in PBS with 0.1% Triton X-100 (Thermo Fisher Scientific, cat. number BP151-500) for 10 min at room temperature and blocked with PBS containing 5% BSA, 5% goat serum (Gibco, cat. number 16210-064) and 0.01% Triton X-100 for 30 min at room temperature. Cells were incubated with IF buffer (PBS, 5% BSA, 0,01% Triton X-100) containing the primary TIA1 antibodies overnight at 4°C. Cells were then washed 3 × 10 min with IF buffer and incubated with corresponding Alexa Fluor 555-conjugated secondary antibodies in IF buffer at a dilution of 1.0 μg/mL for 1 hr at room temperature with DAPI (Thermo Fisher Scientific, cat. Number D3571) fluorescent stain, used to label the nuclei. Cells were washed 3 × 10 min with IF buffer and once with PBS.
Images were acquired on an ImageXpress micro widefield high-content microscopy system (Molecular Devices), using a 20x/0.45 NA air objective lens and scientific CMOS camera (16-bit, 1.97mm field of view), equipped with 395, 475, 555 and 635 nm solid state LED lights (Lumencor Aura III light engine) and bandpass emission filters (432/36 nm, 520/35 nm, 600/37 nm and 692/40 nm) to excite and capture fluorescence emission for DAPI, CellTrackerTM green, Alexa fluor 555 and CellTrackerTM deep red, respectively. Images had pixel sizes of 0.68 x 0.68 microns. Exposure time was set with maximal (relevant) pixel intensity ~80% of dynamic range and verified on multiple wells before acquisition. Since the IF staining varied depending on the primary antibody used, the exposure time was set using the most intensely stained well as reference. Frequently, the focal plane varied slightly within a single field of view. To remedy this issue, a stack of three images per channel was acquired at a z-interval of 4 microns per field and best focus projections were generated during the acquisition (MetaExpress v6.7.1, Molecular Devices). Segmentation was carried out on the projections of CellTrackerTM channels using CellPose v1.0 on green (WT) and far-red (KO) channels, using as parameters the ‘cyto’ model to detect whole cells, and using an estimated diameter tested for each cell type, between 15 and 20 microns.32 Figures were assembled with Adobe Photoshop (version 24.1.2) to adjust contrast then assembled with Adobe Illustrator (version 27.3.1).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)