Keywords
Pseudomonas aeruginosa, BTEX, genes rhlABC, genes mlaABCD, genes pqs
This article is included in the Genomics and Genetics gateway.
Pseudomonas aeruginosa, BTEX, genes rhlABC, genes mlaABCD, genes pqs
Polycyclic aromatic hydrocarbons (PAHs) and BTEX aromatic hydrocarbons (benzene, toluene, ethylbenzene and xylene), widely distributed in the environment in complex mixtures, give us a significant environmental challenge due to their abundance and global recognition as pollutants (Andreoni & Gianfreda, 2007). The World Health Organization has also recognized BTEX as potent carcinogens (Chicca et al., 2020). Petroleum hydrocarbon spills are a serious environmental problem, as they constitute a source of soil and water pollution, disrupting aquatic and terrestrial ecosystems by influencing their structure and biological processes (Shi et al., 2015). Because of their persistence in the environments, they cause harm to human health and to flora and fauna (Dongen, 2000).
In spite of the toxicity risks, hydrocarbon polluted places are the best source to isolate aromatic and aliphatic hydrocarbon degrading microoganisms (Salgado Brito et al., 2008). Among these microorganisms are bacteria belonging to various genera such as Bacillus, Burkholderia, Acinetobacter, Rhodococcus, and Pseudomonas (Ghosal et al., 2016). For instance, strains like Pseudomonas aeruginosa N002, P. aeruginosa SJTD-1, and P. aeruginosa DQ8 have been identified as alkane-degrading organisms, while Massilia aromaticivorans, Pseudomonas putida, Pseudomonas fluorescens, Rhodococcus, Bacillus sp., and Streptomyces sp. have shown potential for BTEX degradation (Kaur et al., 2023). Additionally, Acinetobacter spp., Alcaligenes spp., Acidovorax spp., Janibacter spp., and Rhodococcus spp. have been reported to be capable of degrading PAHs (Shahsavari et al., 2019).
Studies related to the characterization and annotation of genes involved in hydrocarbon degradation are useful to understand the physiological and metabolic processes in the intrinsic dynamics of native microorganisms at the level of molecular biodiversity, genomics, transcriptomics, and metabolomics (Kumari et al., 2023).
In Pseudomonas aeruginosa, it is necessary to consider the quorum sensing hierarchical complex network (García-Reyes et al., 2020; Kuang et al., 2020; J. Lee & Zhang, 2015; Meng et al., 2020): the rhl system (rhlABR operon, rhlC, rhlI, and rhlG genes); las system (lasR, lasI genes) (Kuang et al., 2020; Meng et al., 2020; O’Connor et al., 2022); PQS system (pqsABCBDE operon, pqsH, pqsL, and pqsR or mvfR genes) (Meng et al., 2020; O’Connor et al., 2022; Xiao et al., 2006); and the IQS system (ambBCDE operon, iqsR gene) (J. Lee & Zhang, 2015; O’Connor et al., 2022). Additionally, genes responsible for BTEX tolerance (mlaABCD genes), as well as those involved in alkane degradation (alkB gene) (Cruz et al., 2020), BTEX (tod, tom, tbu, tou, pTOL pathways), and PAHs (TOD, NAH, OCT plasmid genes, among others), have been reported.
Studies of genes related to the degradation of monoaromatic hydrocarbons have been carried in Pseudomonas putida. The toluene-degrading plasmid (TOL) has been studied in P. putida mt-2, which encodes metabolic pathways for the degradation of toluene, m-xylene, and p-xylene into carboxylic acids. The xylN genes of the TOL plasmid encode a m- xylene transporter porin (Kasai et al., 2001). Similarly, the degradation of toluene follows six differentiated metabolic pathways, which involve the tod gene encoding toluene dioxygenase, the tmo/tbm/tbc genes responsible for synthesizing toluene 2-monooxygenase enzyme, tbu/tbh genes encoding toluene-3-monooxygenase, tmoABCDEF genes encoding different subunits of ortho toluene 4-monooxygenase protein, and tbc1FEDCBA/tbc2ABCDEF genes encoding meta toluene 4-monooxygenase (Parales et al., 2008). On the other hand, the genes involved in the aerobic degradation of benzene are the benzA genes encoding an enzyme that carries out the initial ring cleavage oxidation reaction of monoaromatic hydrocarbons, and this enzyme has been studied in Pseudomonas putida AQ8 (Chicca et al., 2020). To date, there are not many records involving strains of the species Pseudomonas aeruginosa in the degradation of monoaromatic hydro- carbons, nor annotated genes associated with this process.
Research on hydrocarbon degradation has extensively explored various species of Pseudomonas, particularly focusing on their ability to break down complex organic compounds in diverse environments. Pseudomonas aeruginosa, renowned for its fluoranthene degradation capability, exhibits robust metabolic capacities. Investigations on this species have spanned multiple domains, including genomics, transcriptomics, metabolomics, and its potential applications in biotechnology (He et al., 2018).
This research is focused in comparing the genetic characterization of genes coding for rhamnolipids production (rhl ) and BTEX degradating genes in six Pseudomonas aeruginosa strains isolated in hydrocarbon polluted environments.
Six strains of Pseudomonas aeruginosa isolated from hydrocarbon contaminated environments, from different geographical sites in Peru (Figure 1) were selected ( Table 1); belonging to the collection of microorganisms of the Laboratory of Microbiology and Microbial Biotechnology (LAMYBIM) of the Faculty of Biological Sciences of the U.N.M.S.M. (Palomino et al., 2017).
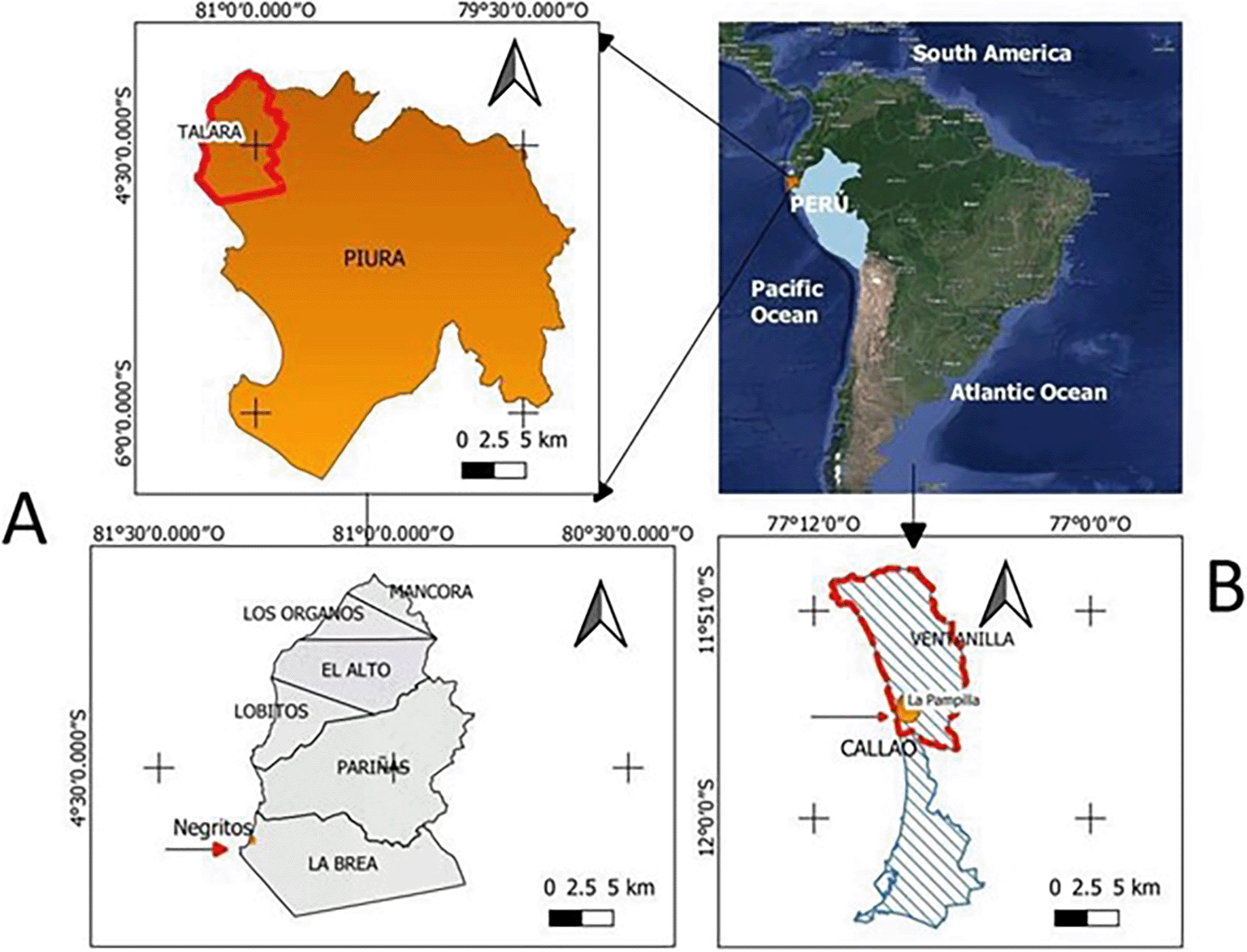
Elaboration: Figure created by the authors as part of this study.
| Selected strains | Codes | Sampling location | Geographic location | Reference |
|---|---|---|---|---|
| Rhmanolipid hyperproducer | 6K-11 | Environments contaminated with hydrocarbons | Talara, Northern Peru | Tabuchi et al., 2015 |
| With emulsifying capacity and heavy metal removal efficiency | PB25 | Environments contaminated with hydrocarbons | La Pampilla, Ventanilla | Giraldo et al., 2014 |
| With Hydrocarbonoclastic capacity | C1BHIC5 | Land treatment field contaminated with crude oil | Talara, Northern Peru | Terán Morales, 2020 |
| With Hydrocarbonoclastic capacity | C3ACETC53a | Land treatment field contaminated with crude oil | Talara, Northern Peru | Terán Morales, 2020 |
| Non-rhamnolipid producer | 2K-1 | Environments contaminated with hydrocarbons | Talara, Northern Peru | Palomino et al., 2017 |
| Non-rhamnolipid producer | 3K-6 | Environments contaminated with hydrocarbons | Talara, Northern Peru | Palomino et al., 2017 |
For the purification process, the strains were subcultured in glass flasks containing 50 mL of Luria Bertani medium, at 37 °C overnight. Subsequently, the methodology of the innuPREP Bacteria DNA kit from analytikjena was followed.
Raw reads were verified with FastQC software (Andrews, S., 2010) and filtered to remove contaminating Illumina adapter sequences and quality trimmed with Trimmomatic v0.36 (Salgado Brito et al., 2008). The resulting filtered reads were assembled using SPAdes v3.14.1 (Ghosal et al., 2016). Assemblies were then filtered to contain only contigs longer than 400 bp. The quality of the assemblies was checked with Quast (Ivanova et al., 2022). For validation assembly, we used Busco (Khan et al., 2001) and mapped reads filtered against the draft genome using Bowtie2 (Zylstra & Gibson, 1989) and Samtools (Furukawa et al., 1993). For annotation, we used the Prokka pipeline, which used Prodigal (Harayama & Rekik, 1993), RNAmmer (Reineke, 1998), Aragorn (Mapelli et al., 2017), and MinCED (Skennerton et al., 2021) to find protein-coding genes, RNA, tRNA, and CRISPR regions, respectively. Finally, we found groups of orthologs using Orthofinder (Cao et al., 2015), adding the proteome of a Pseudomonas aeruginosa strain (GCA_000006765.1) for this analysis.
Trimming quality (Phred Q > 25) and removal of adapters were conducted with Trimmomatic v0.36 (Bolger et al., 2014) and TrimGalore software (Felix Krueger et al., 2023), respectively. De novo assembly was performed using SPAdes v3.10.1 (Bankevich et al., 2012), testing different k-mers (from 23 to 123). SSPACE v2.0 (Boetzer et al., 2011) was used to join contigs to build scaffolds with an iteration of raw read alignment over the contigs to minimize the gaps. A final gap-filling step was performed using GapCloser (Boetzer & Pirovano, 2012) to generate a draft genome. We used QUAST v5.2.0 (Gurevich et al., 2013) for assembly statistics. The completeness and consistency of the assembled genome were estimated using the Benchmarking Universal Single-Copy Orthologs (BUSCO) software (Simão et al., 2015) and CheckM (Parks et al., 2015). These assemblies have been deposited at DDBJ/ENA/GenBank under the accessions: JAOAQG000000000, JAOAQH000000000, JAOAQI000000000, JAOAQJ000000000, JANATH000000000, JANZYL000000000.
The functional annotation of the obtained genomes was carried out using BAKTA v1.6.1 (Schwengers et al., 2021), a precise annotation tool, using a local conda environment (Anaconda v23.5.0). We used BAKTA’s database in its 2022-08-25 release. Standard parameters were used for this functional annotation.
For comparison purposes, Pseudomonas genomes were retrieved from public repositories based on their quality and completeness. All considered genomes of the Pseudomonas aeruginosa species were downloaded from the Reference Sequences Database for comparison (RefSeq; October 2022). Genomes were selected based on the quantity and quality according to the tool QUAST v5.2.0 (Gurevich et al., 2013).
Average Nucleotide Identity (ANI) analysis was performed using pyANI (v. 0.2.12) (Pritchard et al., 2016). A heatmap was generated using the pheatmap package in R (v. 4.1.2). A tblastx was performed using the assembled genomes with a focus in 2 genes of interest, and the genetic diagram was built using the genomes package in R. The phylogenetic reconstruction of multiple Pseudomonas aeruginosa strains was built, leveraging the core gene alignment obtained during pangenome reconstruction through Panaroo (Tonkin-Hill et al., 2020). Subsequently, RAxML v8.2.13 (Stamatakis, 2014) was employed to construct a phylogenetic tree based on the aligned core genes. The model was selected using ModelTest NG (Darriba et al., 2020), with the TVM+I+G4 model chosen as the optimal one for this analysis. The phylogenetic analysis was run with 100 bootstraps. The phylogenetic tree was graphically edited using the iTOL v5 web tool (Letunic & Bork, 2021).
For gene topology and synteny, we performed a tblastx between the contigs of interest. We performed this for 2 genes of interest: rhl and mla. For drawing the topology and the synteny obtained, we used the R package, gggenome (Hackl et al., 2024). For the rhl system, we filtered the tblastx results based on E-value (<0.01), coverage length (≥20) and identity (≥30). For the mla system, we filtered by criteria based on E-value (<0.01), coverage length (≥100) and identity (≥30). PB25’s alignment against P. putida was filtered following more lax criteria (E-value (<0.01), coverage length (≥20) and identity (≥30)).
The obtained genomes presented a very consistent length, with the 2K-1 strain being superior in length compared to the other genomes, with approximately 6 Mbp in size. Most of the other strains had a length of 5.6 Mbp, with C3ACET53a and C1BHIC5 being larger by 0.2 Mbp. The GC content did not vary significantly, with most strains having approximately 66% GC content. The largest N50 value was observed for C1BHIC5 ( Table 2).
The Average Nucleotide Identity (ANI) was compared between various strains of Pseudomonas aeruginosa using the algorithm MUMMER. The sequenced strains presented a notable similarity when compared to other Pseudomonas aeruginosa strains, verifying that our strains belong to the Pseudomonas aeruginosa species (>0.95) (Jain et al., 2018). The lowest identity score was 97.5% (0.975), while the highest score was 99.9% (0.999) ( Figure 2).
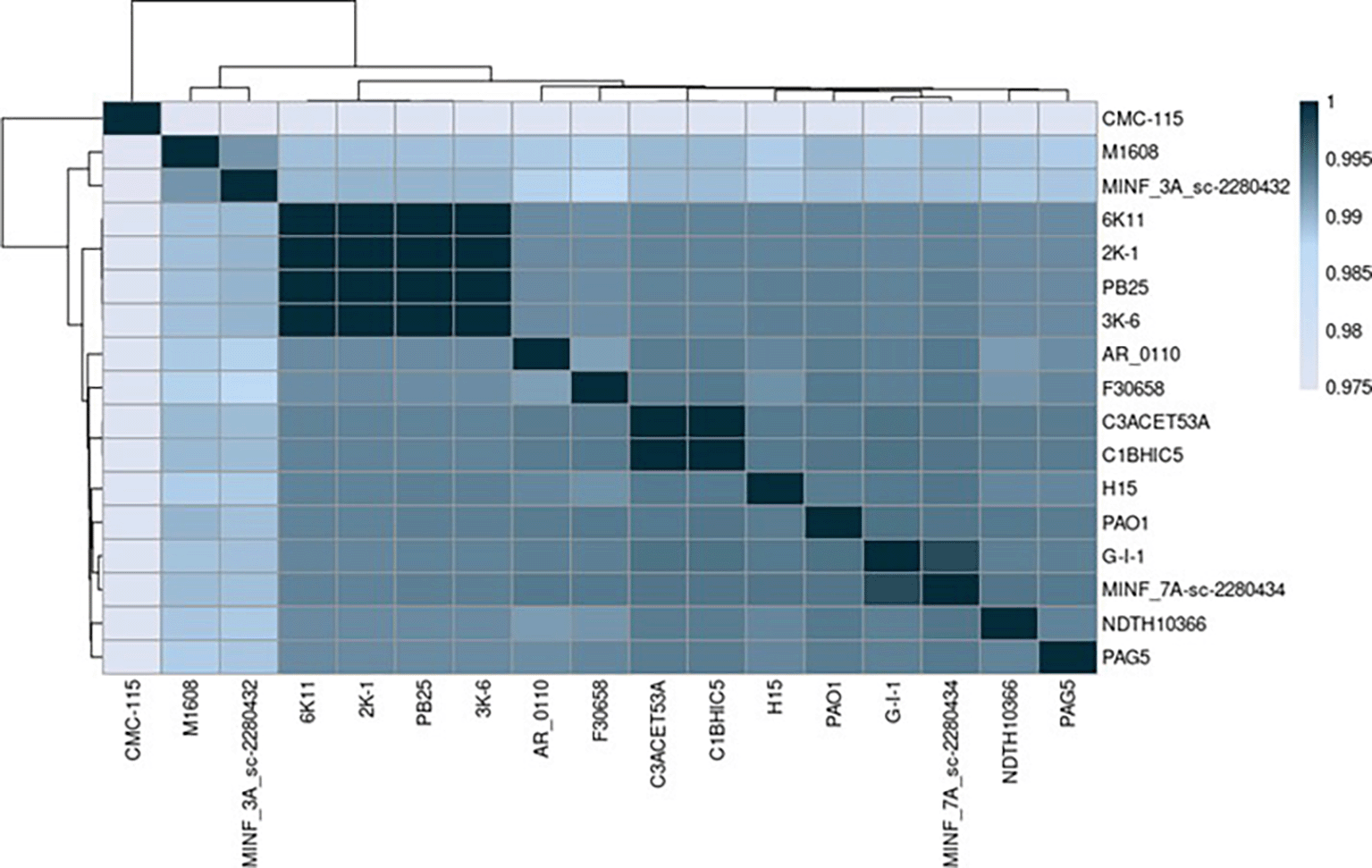
Elaboration: Figure created by the authors as part of this study.
A small pangenome is represented in Figure 3A, showing a comparison between the isolated strains, PAO1, and PA7. This comparison illustrates how the strains described in this work diverge from some representative strains of Pseudomonas aeruginosa. Among these 8 strains, 4068 genes are shared. Of these, 65 genes are common to all isolated strains, excluding PA7 and PAO1. Additionally, 108 genes are exclusively shared by C3ACET53A and C1BHIC5, while 197 genes are exclusively shared by the group comprising 6K-11, PB25, 3K-6, and 2K-1. This indicates certain similarities and shared genes among the isolates of interest, as well as a clear separation between the isolated strains into two distinct groups.

Elaboration: Figure created by the authors as part of this study.
A total of 3544 genes were classified as core genes in a pangenome built with 93 strain, including the ones isolated for this study. These core genes are likely to be involved in essential cellular functions and housekeeping processes. On the other hand, 11814 genes were categorized as accessory or variable genes, reflecting their presence in only a subset of the genomes. These genes were classified in 3 categories: Softcore, shell and cloud genes, depending on their presence in the pangenome. These accessory genes contribute to the unique genetic diversity of each strain and may play crucial roles in shaping their distinct phenotypic characteristics ( Table 3).
A phylogenetic tree was constructed based on various strains previously reported in other studies. Bootstrap values reached a value of 100 for most cases. The branch length for PA7 had to be reduced because of its unusual divergence (original branch length = 9.9999513099). We attempted to reconstruct the Pseudomonas aeruginosa “biotypes” found by Palleroni et al. (1973). We determined that all six sequenced strains belong to Clade 1, in which there are also strains such as the reference strain PAO1. However, these strains are divided in 2 groups that are distant from each other in Clade 1. The 6K11, 2K-1, PB25 and 3K-6 strains are situated in a clade shared with strains T38079, F9670, S86968, ATCC 27853, PA96 and M18. Meanwhile, strains C1BHIC5 and C3ACET53A share a clade with strains 8380 and SJTD-1 ( Figure 4).
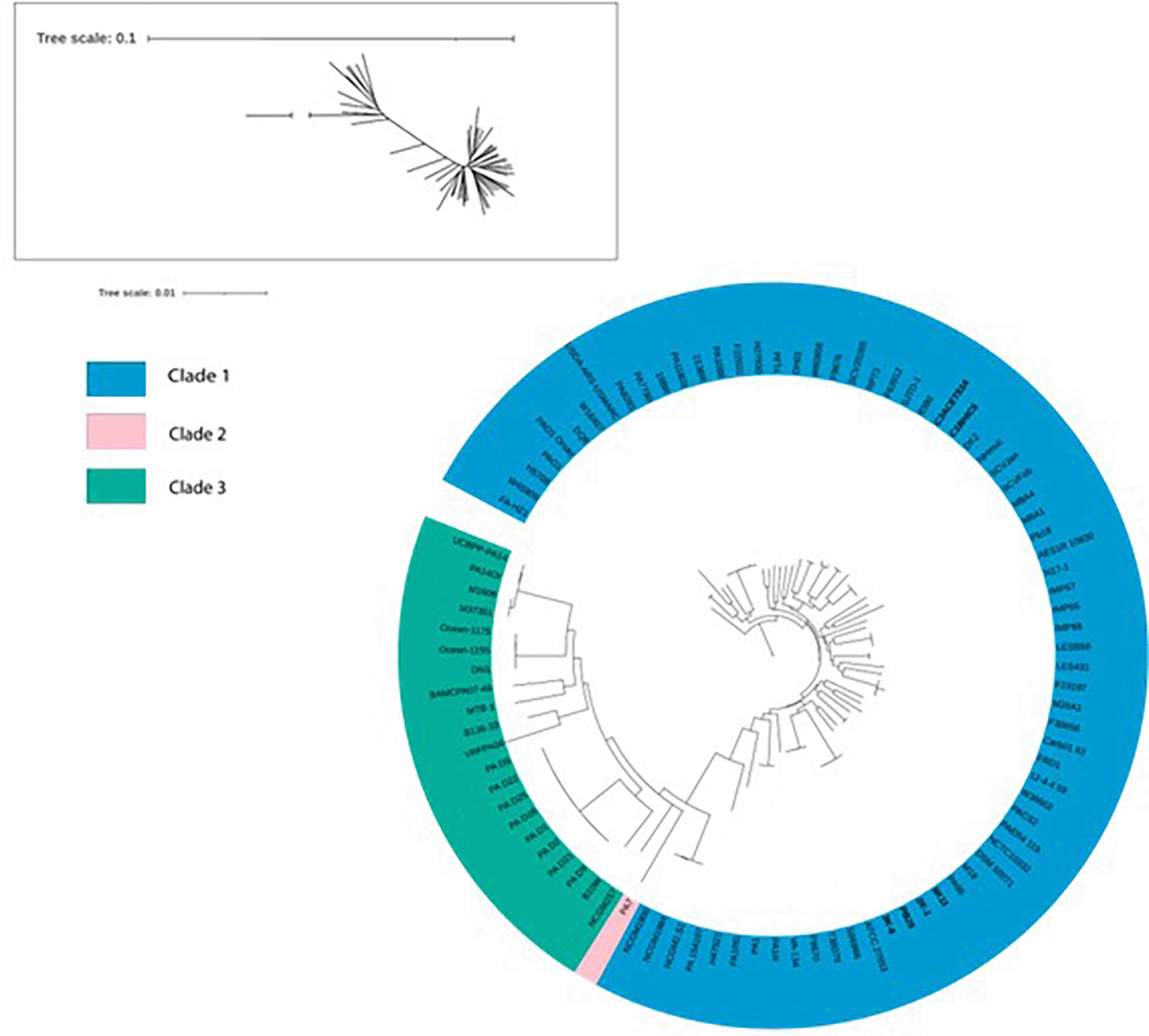
In bold letters, the isolates of interest.
Elaboration: Figure created by the authors as part of this study.
The rhl gene family, primarily found in Pseudomonas aeruginosa and Burkholderia species, is responsible for the biosynthesis of rhamnolipids. Rhamnolipids are surface-active molecules composed of rhamnose and β-hydroxydecanoic acid. These biosurfactants have been thoroughly investigated due to their involvement in various multicellular behaviors.
We evaluated the organization and synteny of rhl genes in several bacterial strains (Figure 6). Our strains presented a double copy of the rhlB gene, except for the C1BHIC5 strain, with only 1 rhlB gene. The rhl genes did not exhibit a radically different organization across all evaluated strains: The genes rhlABRI are always neighboring each other. They are positioned in a 3′-5′ sense, except in C3ACET53A. This sense is the one exhibited in the PAO1 strain too. The rhlG gene does not have the same position in all strains. It always appears neighboring the rhlC gene. All rhlG genes in the strains have a 5′-3′ orientation, except in C3ACET53A. The rhlC gene, likewise, appears in different positions, but always neighboring the rhlG gene. Its orientation varies between strains. The rhlB gene is present in 2 positions in the genome. One copy is part of the rhlABRI cluster, while the other is situated in another position, far from the cluster. The rhlABRI cluster is also part of the PAO1 genome.
Furthermore, we observed a significant synteny between the rhlB gene in the recovered isolates. The copy situated far from the rhlABRI presents a clear synteny between all the strains, and seems to closely relate between each of these strains ( Table 3).
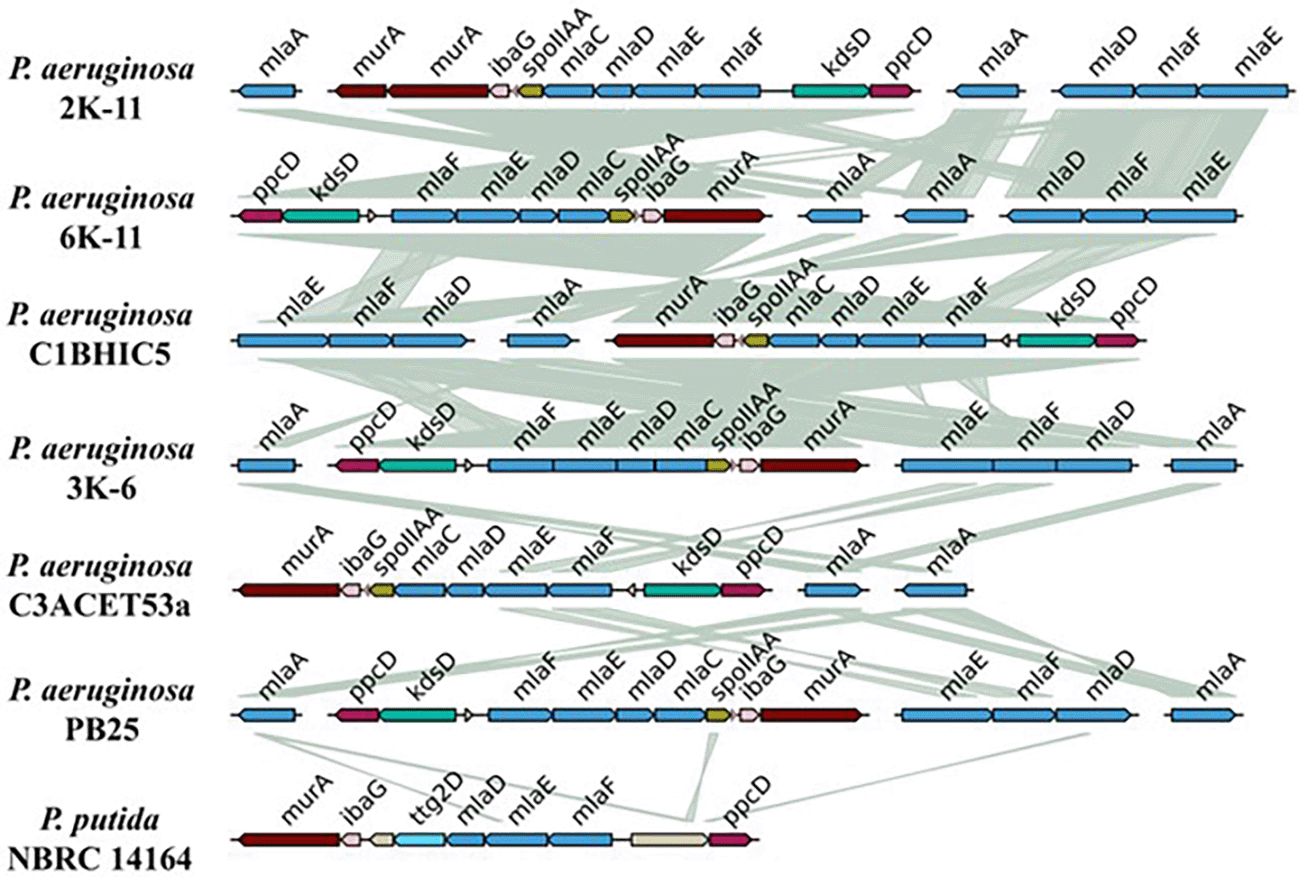
Due to the toluene tolerance previously shown by the strains (Querebalú García, 2020, data not available), data no published), the mla system was sought as a gene operon of interest. Genomically, this system was present with a similar structure in all the isolated P. aeruginosa strains ( Figure 7). Almost all the strains exhibited two copies of the mlaA gene, except for the strain C1BHIC5, which presented only 1 copy of this gene. The mlaFEDC cluster is present in all strains. On the other hand, the mlaEFD cluster is present in all strains except for the strain C3ACET53A. The orientation of these clusters varies among the sequenced strains. Regarding the synteny between these clusters, it is clear that there was high synteny between these strains, and the mla gene blocks were highly similar. Interestingly, when compared with Pseudomonas putida, this species presented the mla genes between the murA and ppcD genes, in a similar conformation to other P. aeruginosa strains. Notably, the gene responsible for toluene tolerance, ttg2D, was also found among these genes in P. putida.
Elaboration: Figure created by the authors as part of this study.

Elaboration: Figure created by the authors as part of this study.
Elaboration: Figure created by the authors as part of this study.
Several studies seek to evaluate the bioremediation potential of Pseudomonas aeruginosa against hydrocarbons, such as Baig et al. (2022), who isolated 2 strains: BAA-427 and ATCC-27853, from oil-contaminated soils (Aslam Refinery, Pakistan); Liu et al. (2022) also isolated P. aeruginosa AQNU-1 in water samples from a lake wetland near a petrochemical industry (Anqing City, China) and Mahjoubi et al. (2021) isolated a halotolerant strain of P. aeruginosa UN14 in hydrocarbon-contaminated sediment (refinery on the coast of the port of Bizerte, northern Tunisia). In Peru, the Laboratory of Microbiology and Microbial Biotechnology of the U.N.M.S.M. has actively isolated strains of Pseudomonas aeruginosa from polluted environments, which present efficient physiological capacity in relation to hydrocarbon degradation. Of interest are strains with hyper-producing capacity of rhamnolipids (Pseudomonas aeruginosa 6K-11); with emulsifying capacity and efficiency for the removal of heavy metals (Pseudomonas aeruginosa PB25) and with hydrocarbonoclastic activity (Pseudomonas aeruginosa C1BHIC5 and Pseudomonas aeruginosa C3ACETC53a).
The species phylogeny was constructed using strains previously reported in various studies, focusing especially on the “biotypes” identified by Palleroni et al. (1973). Our strains, originated from environmental sources, formed 2 distinct groups within Clade 1, along with clinical origin strains. Unlike other pathogenic bacteria, genomic studies have shown that clinical and environmental isolates of P. aeruginosa do not exhibit particular genomic differences, confirming that genetic variability related to the environment is reduced in this species (Grosso-Becerra et al., 2014). This would explain why the origin of the strains did not determine the clade they formed in the phylogeny of P. aeruginosa. Likewise, it would also explain the results observed in the ANI matrix, in which the percentages are not very different from each other.
The pangenomic comparison also showed a large number of accessory genes (n=10,397; 67%) and a reduced core genome (n=3,544; 23%). This coincides with previous pangenomic studies that have highlighted the importance of horizontal gene transfer in this species (Freschi et al., 2019).
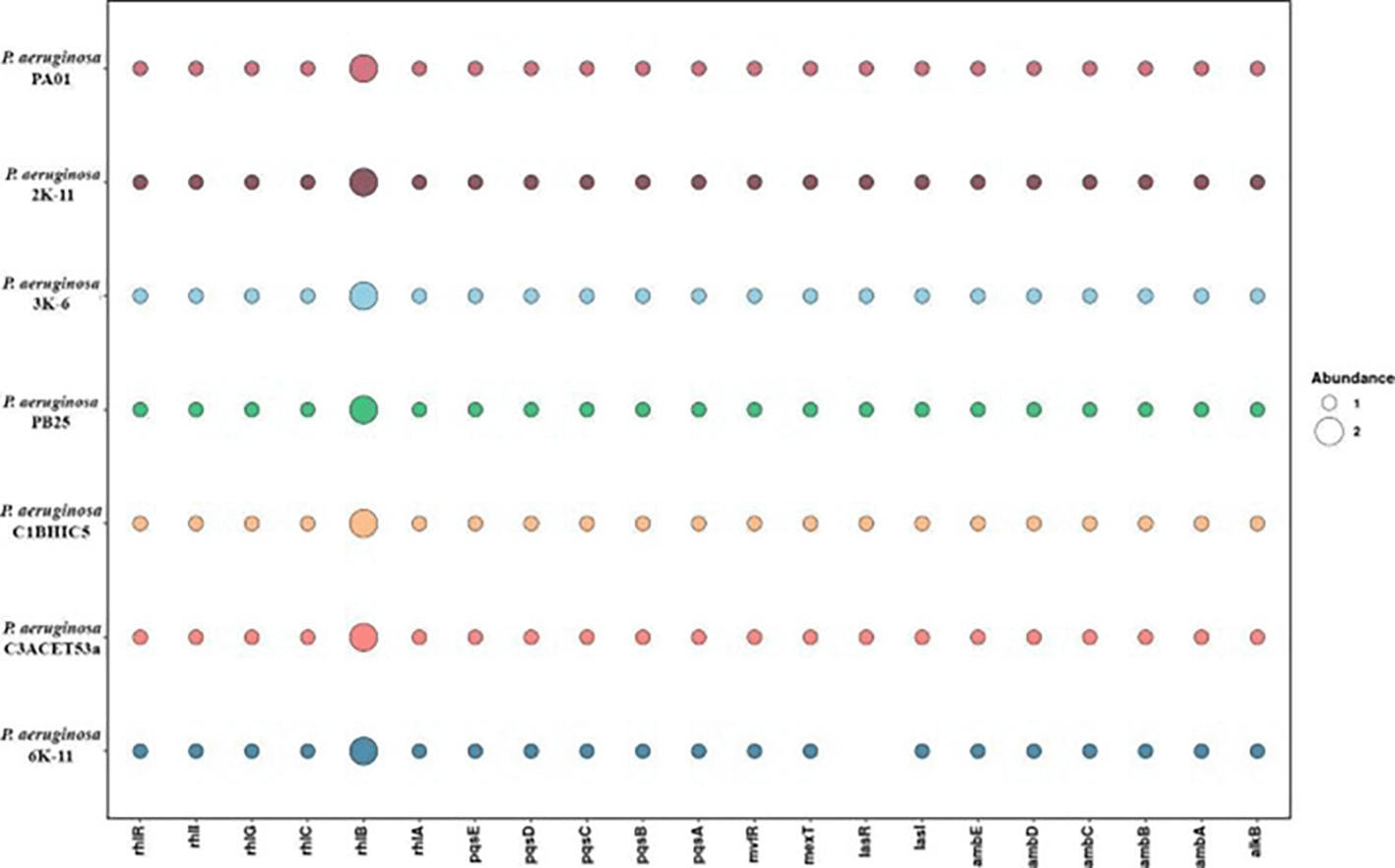
Pseudomonas aeruginosa utilizes a quorum sensing (QS) system to regulate the synthesis of rhamnolipids (RLs), which are essential for the emulsification and assimilation of hydrocarbons (Xu et al., 2020). The rhlABC genes are responsible for the production of these RLs in Pseudomonas aeruginosa strains isolated from petroleum-contaminated environments (Castro et al., 2022; Ji et al., 2016). These genes encode the enzymes RhlA, RhlB, and RhlC, which transform precursors into mono-rhamnolipid and di-rhamnolipid. We have evidenced the presence of these genes in all strains analyzed through comparative genomic studies ( Figure 5).
Regarding quorum sensing interactions, the las system regulates the expression of virulence genes and activates the pqs and rhl systems (Lee & Zhang, 2015), while the pqs system regulates more than 35 loci related to virulence (Li et al., 2022). The rhl system controls both virulence genes and rhamnolipid production (García-Reyes et al., 2020). In our analysis of these genes across several strains, we found that all strains have a similar gene abundance. However, the 6K-11 strain stands out due to the absence of the lasR gene (Figure 7). These findings regarding the absence or presence of these genes could be due to the respective virulence mechanisms in each strain (Dötsch et al., 2015).
In terms of the degradation of BTEX hydrocarbons (benzene, toluene, ethylbenzene, and xylene), specific genes responsible for this process have already been identified in various microorganisms (Choi et al., 2013; Lee et al., 2019; Lin et al., 2002), primarily studied in species of Pseudomonas and P. putida (Bacosa et al., 2021). In our study, in an attempt to corroborate hydrocarbonoclastic physiological activity, we searched for these genes within the complete genome but were unsuccessful in all the strains analyzed. Therefore, we searched for orthologous genes from the ttg2 operon previously found in Pseudomonas putida, initially linked to toluene tolerance (Yero et al., 2021), with this system being the mla. MlaA removes glycerophospholipids from the outer leaflet of the outer membrane (OM) and deliver them to the MlaFEDB complex in the inner membrane (IM) via the periplasmic substrate-binding protein MlaC (Kaur et al., 2023).
The genetic organization of the mla system in the synteny showed discontinuity, as the mlaCDEF genes are grouped in an operon, while mlaA is separated in the genome. This aligns with what was reported by Kaur and Mingeot -Leclercq (2024), who mentioned that in gram-negative bacteria, including P. aeruginosa, the system is organized in this way. However, the mlaB gene was not found in the operon, consistent with the absence of this gene in alpha-proteobacteria and epsilon-proteobacteria.
Benzene, toluene, ethylbenzene, and xylene, collectively known as BTEX, are common pollutants of soils and groundwater. BTEX and other polycyclic aromatic hydrocarbons are extremely toxic to microorganisms due to their accumulation in hydrophobic cell membranes (Choi et al., 2013; Hąc-Wydro et al., 2019). Therefore, the genes studied may be involved in resistance to BTEX. Pseudomonas aeruginosa possesses a complex genetic and enzymatic regulatory system that allows it not only to produce rhamnolipids essential for the emulsification and assimilation of hydrocarbons but also to tolerate aromatic compounds such as BTEX
Studies on genes related to the degradation of monoaromatic hydrocarbons have been conducted in Pseudomonas putida. The toluene-degrading plasmid (TOL) has been studied in P. putida mt-2, which encodes metabolic pathways for the degradation of toluene, m-xylene, and p-xylene into carboxylic acids. The xylN genes of the TOL plasmid encode a porin transporter for m-xylene (Kasai et al., 2001). Similarly, toluene degradation follows six differentiated metabolic pathways, involving the tod gene, which encodes the enzyme toluene dioxygenase, the tmo/tbm/tbc genes responsible for the synthesis of the Toluene 2-monooxygenase enzyme, tbu/tbh genes that encode Toluene 3-monooxygenase, tmoABCDEF genes that encode different subunits of the ortho Toluene 4-monooxygenase protein, and tbc1FEDCBA/tbc2ABCDEF genes that encode meta Toluene 4-monooxygenase (Parales et al., 2008). On the other hand, the genes involved in the aerobic degradation of benzene are the benzA genes, which encode an enzyme that carries out the initial oxidation reaction of the ring-cleavage of monoaromatic hydrocarbons, and this enzyme has been studied in Pseudomonas putida AQ8 (Chicca et al., 2020). To date, there are not many records involving strains of the species Pseudomonas aeruginosa in the degradation of monoaromatic hydrocarbons, nor annotated genes associated with this process.
If you are using Overleaf, either select “Submit” then F1000Research, or click “Submit to F1000Research” in the top right-hand corner. Alternatively, generate a PDF file of your project and submit this alongside a zip file containing all project files (includes the source files, style files, and PDF) using online submission form.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)