Keywords
peptidomelanin; melanin; peptides; ecology
This article is included in the Cell & Molecular Biology gateway.
peptidomelanin; melanin; peptides; ecology
Melanin comprises an ancient, diverse class of high–molecular-weight, dark aromatic biopolymers4 found across all kingdoms of life.5,6 They enhance survival under stresses such as ionizing radiation,7 oxidative stress,8 and metal toxicity.1 Despite their ecological9 and pathogenic significance,10 melanins remain poorly structurally resolved due to insolubility, chemical heterogeneity, and resistance to degradation.11 The best-characterized fungal melanins derive from L-3,4-dihydroxyphenylalanine (L-DOPA)12 and 1,8-dihydroxynaphthalene (DHN),13 and are typically tightly associated with the cell wall, limiting characterization.11
Soluble forms of melanin are less common but have been reported. Pyomelanin is a nitrogen-free, water-soluble polymer derived from homogentisic acid during tyrosine catabolism.14 Its lower molecular weight and carboxyl groups enhance water solubility. Soluble melanin from Glyomastix polychroma forms small nanoparticles that remain colloidally suspended in water.15 Soluble melanins from several microbes have also been reported, though their detailed biochemical composition is usually not characterized.16–21
Our laboratory previously discovered and characterized peptidomelanin ( Figure 1A,B), a soluble melanin composed of an L-DOPA-derived phenolic core covalently linked to short heterogeneous peptides forming a solubilizing “corona”.1 It is synthesized via the Srivatsan pathway,2,22 in which a broad-spectrum copper oxidase (BroSCO) copolymerizes cysteinylated peptides with the phenolic core. Peptidomelanin can also be polymerized in vitro and functionalized with synthetic peptides.3
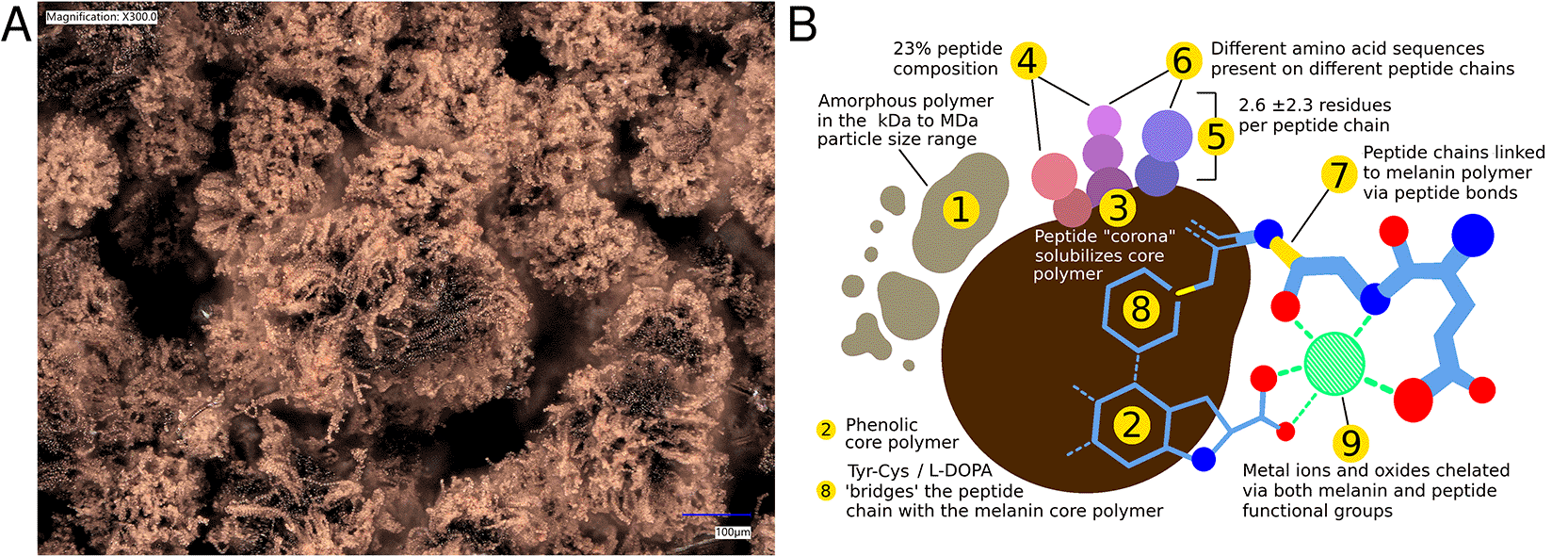
(B) Our model of the biochemical composition of peptidomelanin. Inferences 1–9 are supported by experiments reported in our previous work.1,2,3 Panel B has been reprinted (adapted or reprinted in part) with permission from Kolipakala, Rakshita Sukruth, et al. “Fungal peptidomelanin: a novel biopolymer for the chelation of heavy metals.” ACS omega 9.34 (2024): 36353–36370. Copyright 2024, the authors.
We previously showed that Peptidomelanin is secreted into the extracellular medium by germinating spores of Aspergillus niger melanoliber (MTCC 13366),1 though the trigger for secretion was unknown. Here, we show that secretion is specifically induced by extracellular peptides, but not amino acids, sugars, or proteins. Fluorescence microscopy reveals peptide binding to the spore surface, suggesting surface-associated sensing that initiates peptidomelanin secretion. These findings provide a basis for future mechanistic studies of the peptidomelanin secretory pathway.
Aspergillus niger melanoliber (MTCC 13366) and Aspergillus niger (MTCC 281) were cultured on Sabouraud dextrose agar (Sisco Research Laboratories Pvt. Ltd. (SRL) code no. 24835) plates supplemented with 1% tyrosine (Sisco Research Laboratories Pvt. Ltd. code no. 18917) and incubated at 37 °C till maturation. Cultures were grown under identical conditions to ensure experimental comparability between strains. Fresh spores were aseptically harvested from mature colonies using a sterile nichrome loop and used immediately for the following experiments.
Our laboratory isolated and identified Aspergillus niger melanoliber from a soil sample collected from Mumbai, India.1 The strain was deposited into the Microbial Type Culture Collection (MTCC, Chandigarh) by the corresponding author (Deepesh Nagarajan) and was made publicly available under the accession number 13366, prior to the start of this work.
Peptidomelanin secretion was assessed by monitoring absorbance of the extracellular medium at 420 nm (OD420) using a colorimeter (Systronics, Digital Photoelectric Colorimeter Type: 112). For each replicate, mature spores of Aspergillus niger melanoliber (MTCC 13366) or Aspergillus niger (MTCC 281), grown on solid media as described previously, were harvested using a sterile nichrome loop, and inoculated into 5 mL of Sabouraud dextrose broth (Sisco Research Laboratories Pvt. Ltd. (SRL) code no. 24835) until the OD420 reached ∼0.6. This solution was diluted using 10 mL of Sabouraud dextrose broth. 2 mL of this diluted solution was pipetted into a borosilicate glass colorimetric cuvette. We tested ≥4 replicates for each fungal strain. OD420 measurements were recorded immediately using timepoints of t=0, 10, 20, 30, 40, 50, 90, 120, 180 and 240 minutes. Cultures were incubated at 37 °C in between measurements. After acquiring data, fold-change values for absorbance were calculated using Equation 1. Results are depicted in Figure 2A.
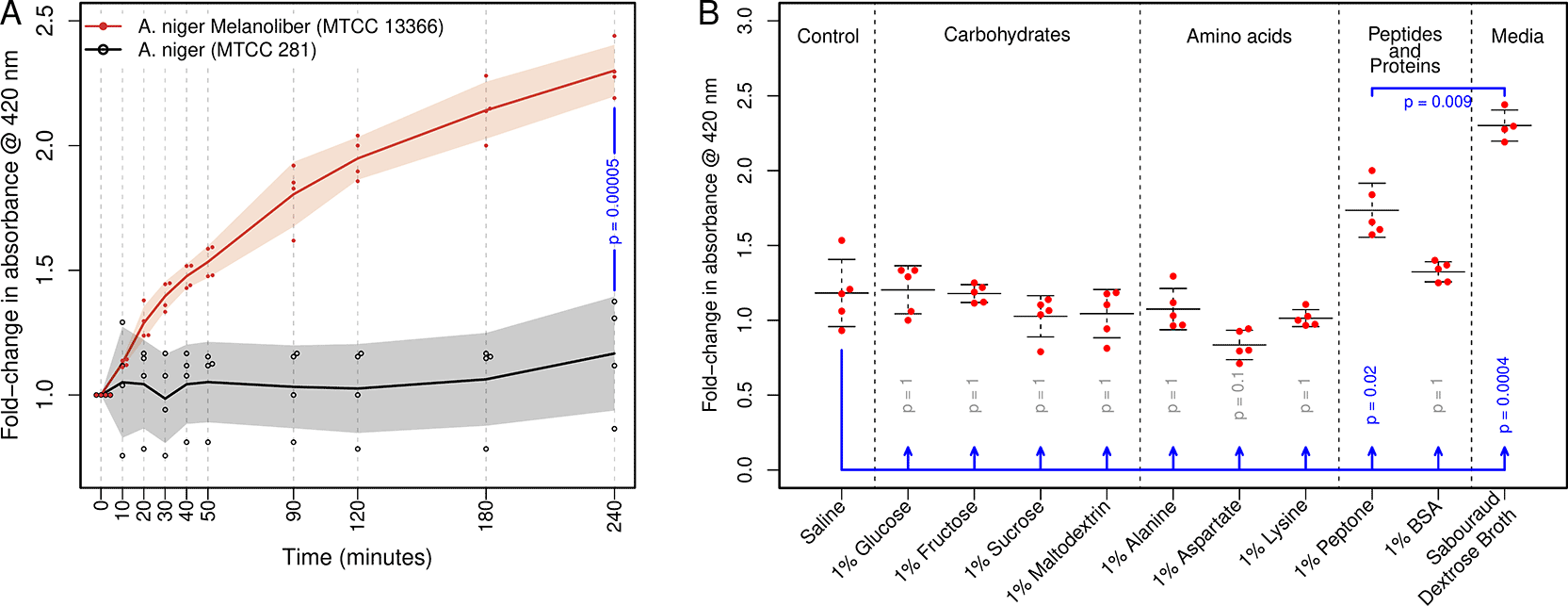
Germinating spores of the type strain of A. niger MTCC 281 do not secrete soluble melanin under the same conditions (p = 0.00005). Melanin secretion was tracked using absorbance at 420 nm. Peptidomelanin secretion from A. niger melanoliber spores is triggered by 1% peptone and Sabouraud dextrose broth. Both media trigger the secretion of significantly more peptidomelanin (p = 0.02 and p = 0.0004, respectively) compared to a 1% saline control condition. All p-values were calculated in R using Student’s t-test. P-values were adjusted for multiple comparisons using the Holm–Bonferroni correction to control the family-wise error rate. Raw data for this image has been made available (refer data availability section).
We prepared 50 mL of 1% (w/v) solutions of glucose (NICE Chemicals (P) Ltd., sucrose from HiMEDIA Laboratories Pvt. Ltd., code no. PCT0607), fructose (Sisco Research Laboratories Pvt. Ltd. (SRL) code no. 42868), maltodextrin (Sisco Research Laboratories Pvt. Ltd. (SRL), code no. 73638), alanine (L-Amino acids kit from HiMEDIA Laboratories Pvt. Ltd. code no. GRM4020), aspartate (L-Amino acids kit from HiMEDIA Laboratories Pvt. Ltd., code no. GRM4020), lysine (L-Amino acids kit from HiMEDIA Laboratories Pvt. Ltd. code no. GRM4020), peptone (HiMEDIA Laboratories Pvt. Ltd., code no. RM001), and bovine serum albumin (BSA) (NICE Chemicals Pvt. Ltd. code no. A31501) in deionized water (Milli-Q). We also prepared saline (1% NaCl, Qualigens Thermo Fisher Scientific India Pvt. Ltd., code no. 7647-14-5) in deionized water, (Milli-Q) as a negative control. Each solution was filter sterilized (0.2 μm, PVDF membrane) before inoculation. For each replicate of each solution, we harvested mature spores of Aspergillus niger melanoliber (MTCC 13366), grown on solid media as described previously, using a sterile nichrome loop. We inoculated the strain into 5 mL of a given solution until the OD420 reached ∼0.6. The given solution was diluted using 10 mL of itself. 2 mL of this diluted solution was pipetted into a borosilicate glass cuvette. We tested 5 replicates for each solution. OD420 measurements were recorded at t = 0 and t = 240 minutes. After acquiring data, fold-change values for absorbance were calculated using Equation 1. Results are depicted in Figure 2B.
Dansylation of bovine serum albumin (BSA) was performed to generate fluorescently labeled peptide substrates. Briefly, 10 mg of BSA was dissolved in 200 μL of 0.4 M sodium bicarbonate (Reachem Laboratory Chemicals Pvt. Ltd., code no. So5720) buffer prepared in deionized water (Milli-Q). In parallel, 10 mg of dansyl chloride (dansyl-Cl, Sigma-Aldrich, code no. 102564199) was dissolved in 200 μL of acetone (Thermo Fisher Scientific India Pvt. Ltd., code no. 67-64-1). The dansyl chloride solution was added to the BSA solution and mixed thoroughly. The reaction mixture was incubated for two hours at 37 °C in the absence of light and covered using aluminum foil.
Following incubation, the reaction mixture was subjected to dialysis against 2 × 1L of deionized water (Milli-Q) to remove unreacted dansyl-Cl. Successful removal of unbound fluorophore following dialysis was visually confirmed by the absence of fluorescence in the dialysate ( Figure 3B, leftmost cuvette). The resulting dansylated BSA solution was then divided into two equal aliquots. One aliquot was retained as intact dansylated BSA, serving as a reference sample ( Figure 3B, middle cuvette). The second aliquot was treated with proteinase K (Sisco Research Laboratories Pvt. Ltd. (SRL), code no. 49936) to generate peptide fragments ( Figure 3B, right cuvette). The proteinase K–treated sample was incubated at 37 °C for 4 hours, after which the enzymatic reaction was terminated by heating at 90 °C for 5 minutes. The resulting dansylated BSA-derived peptides were used for subsequent fluorescence microscopy experiments.
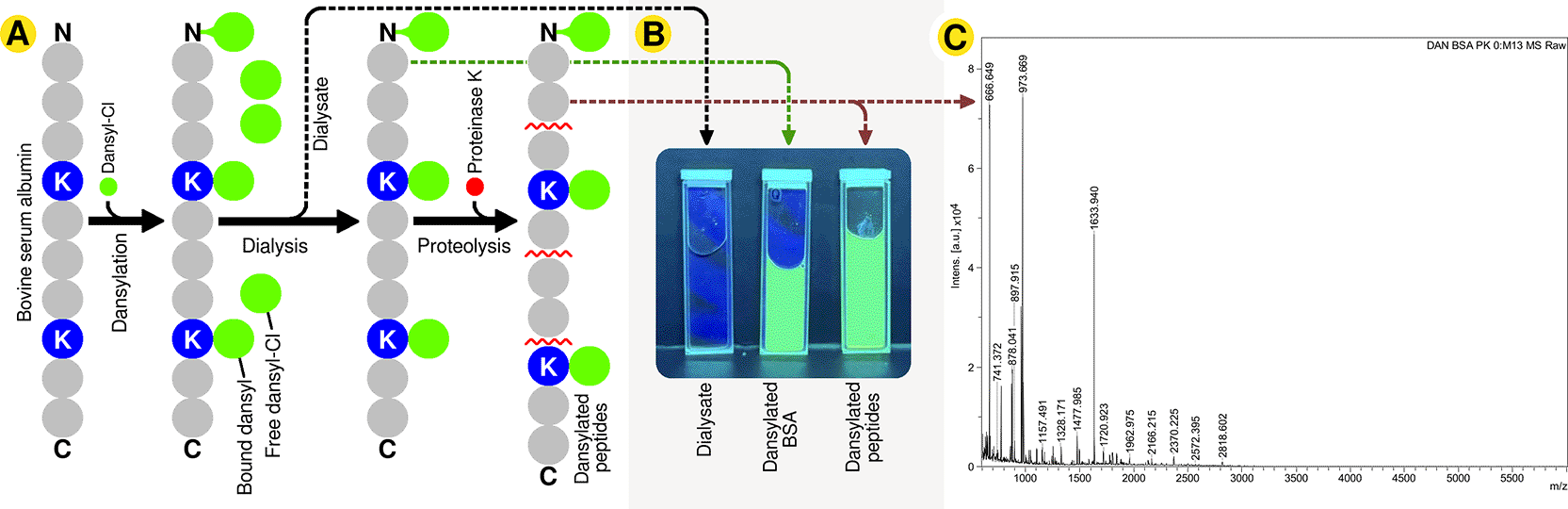
(A) Dansyl-Cl was reacted with bovine serum albumin (BSA), followed by dialysis to remove unbound dansyl-Cl and proteolysis with Proteinase K to generate short dansylated peptides. (B) The dialysate after dansylation showed no fluorescence, confirming removal of unbound dansyl-Cl, while dansylated BSA and the resulting peptides remained fluorescent after dialysis and proteolysis. (C) Matrix-Assisted Laser Desorption/Ionization mass spectrometry (MALDI) spectrum of proteinase K–treated dansylated BSA peptides showing multiple low m/z peaks corresponding to proteolytic fragments.
Scanning electron microscopy (SEM) analyses were conducted using a FEI (Field Electron and Ion Company) Quanta 200 scanning electron microscope at Icon Laboratories Pvt. Ltd., Mumbai. The specimens were examined under low-vacuum conditions at an accelerating voltage of 20 kV, with the chamber pressure maintained at 65 Pa. SEM micrographs are presented in Figures 4A,D.
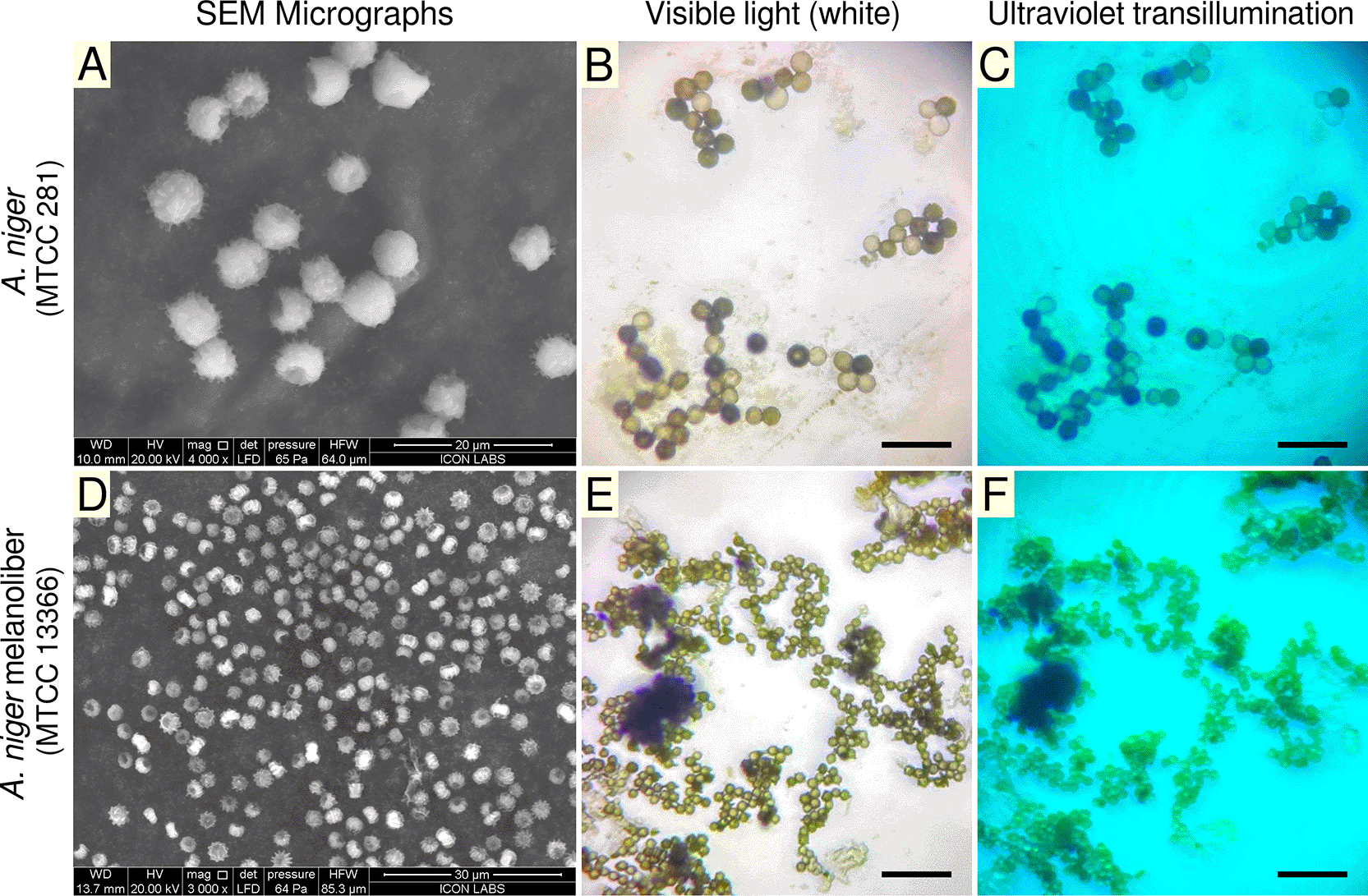
(A) Scanning electron micrograph of Aspergillus niger (MTCC 281) type strain spores, which are larger and show randomly distributed echinulations. (B) Light micrograph of MTCC 281 after treatment with dansylated peptides. (C) UV transillumination of MTCC 281 spores showing no fluorescence. (D) Scanning electron micrograph of A. niger melanoliber (MTCC 13366) spores, which are smaller and display longitudinal striations. (E) Light micrograph of MTCC 13366 after treatment with dansylated peptides. (F) UV transillumination of MTCC 13366 spores showing fluorescence, indicating peptide binding. Scale bar (B, C, D, F): 20 μm.
Mature spores of Aspergillus niger (MTCC 281) and Aspergillus niger melanoliber (MTCC 13366) were assayed for peptide binding. For each strain, mature colonies were grown as described previously (subsection: Fungal strains and culture conditions). 100 μL of dansylated peptides (derived from bovine serum albumin; subsection: Dansylation of BSA and cleavage using Proteinase K) was placed on a concave glass slide. One loopful of spores from a mature colony was mixed with the peptide solution and immediately visualized under bright field using a CxL MONO compound microscope (Labomed Inc.), with a 40× objective and 10× eyepiece ( Figures 3B,E). Samples were then irradiated and visualized using UV-C transillumination from a Philips UV-Stick UVC disinfection wand positioned between the condenser and stage to excite the dansyl fluorophore ( Figures 3C,F).
Proteinase K–treated, dansylated bovine serum albumin (BSA) was submitted to the LC–MS facility at the Indian Institute of Science, Bangalore, for MALDI mass spectrometry. Sample preparation, matrix selection, instrument calibration, and data acquisition were performed by facility staff using a Rapiflex MALDI Tissuetyper (Bruker Daltonics) operated in positive-ion reflector mode. The resulting spectra were provided to the authors for analysis and interpretation ( Figure 3C).
Spores from A. niger melanoliber (MTCC 13366), our peptidomelanin-secreting strain, and A. niger (MTCC 281), a type strain, were incubated in Sabouraud dextrose broth. These cultures were incubated at 37 °C for 4 hours under static conditions ( Figure 2A), and the absorbance (420 nm) of the extacellular media was tracked to determine peptidomelanin secretion. A. niger melanoliber (MTCC 13366) rapidly secreted peptidomelanin within the first 90 minutes, followed by a sustained rise up to 4 hours, whereas Aspergillus niger (MTCC 281) showed negligible secretion across all measured time points. These findings demonstrate that peptidomelanin secretion is restricted to Aspergillus niger melanoliber (MTCC 13366), and does not represent a conserved or generalizable trait across the Aspergillus genus.
We further tested the ability of different classes of molecules to trigger peptidomelanin secretion from germinating spores of Aspergillus niger melanoliber (MTCC 13366) ( Figure 2). All conditions were incubated at 37°C for 4 hours, with peptidomelanin secretion into the extracellular medium monitored via absorbance at 420 nm. All conditions were compared to a saline control (1% NaCl) for statistical significance testing. Our findings are presented below in enumerated form:
1. Carbohydrates alone do not trigger peptidomelanin secretion. Spores incubated in 1% solutions of glucose (monosaccharide), fructose (monosaccharide), sucrose (disaccharide), and maltodextrin (oligosaccharide) displayed no significant increase in peptidomelanin secretion.
2. Amino acids did not trigger peptidomelanin secertion. Spores incubated in 1% solutions of alanine (neutral), aspartate (negatively charged), and lysine (positively charged) displayed no significant increase in peptidomelanin secretion.
3. Peptides, but not proteins, triggered peptidomelanin secretion. Spores incubated in 1% peptone released significantly more peptidomelain into the extracellular medium (p = 0.02). However, spores incubated in 1% bovine serum albumin (BSA) did not display a significant increase in peptidomelanin secretion.
4. Sabouraud dextrose broth consists of 1% peptone and 4% glucose. Spores incubated in this medium released significantly more peptidomelanin, compared to both the saline control (p = 0.0004) and the 1% peptone condition (p = 0.009). We hypothesize that glucose acts as a secretory adjuvant. While glucose alone does not trigger any significant increase in peptidomelanin secretion, in combination with peptides it triggers more peptidomelanin secretion.
It stands to reason that germinating spores of A. niger melanoliber (MTCC 13366) would recognize extracellular peptides via peptide-spore binding, and that such binding would trigger peptidomelanin secretion. We tested this hypothesis using dansylated bovine serum albumin- (BSA-) derived peptides. Dansyl chloride (Dansyl-Cl) is a fluorophore possessing an excitation peak at 335 nm (ultraviolet) and an emission peak at 518 nm (green). The dansyl group covalently links to primary amines, such as the N-terminal of proteins or lysine side-chains. In short, BSA was dansylated, dialyzed to remove excess unreacted dansyl-Cl, and cleaved using proteinase K to produce short dansylated peptides ( Figure 3A). Both dansylated BSA and dansylated, proteinase K-cleaved BSA remained fluorescent under UV illumination (Figure 3B), confirming successful dansylation. Dansylated BSA-derived peptides possessed masses ranging from ∼0.6 kDa to ∼2 kDa, as determined by Matrix-Assisted Laser Desorption/Ionization (MALDI) mass spectrometry. This confirms successful proteinase K cleavage.
Morphologically, spores of the type strain of A. niger (MTCC 281) are relatively large, and possess evenly distributed surface echinulations ( Figure 4A). When treated with dansylated peptides, spores of A. niger (MTCC 281) do not display fluorescence under visible light ( Figure 4B) or ultraviolet transillumination ( Figure 4C).
Spores of A. niger melanoliber (MTCC 13366) are relatively small and possess longitudinal striations originating from the polar regions ( Figure 4D). When treated with dansylated peptides, spores of A. niger melanoliber (MTCC 13366) display weak fluorescence under visible light ( Figure 4E, weak green), and strong fluorescence under ultraviolet transillumination ( Figure 4F, bright green). These observations confirm that dansylated peptides selectively bind to spores of A. niger melanoliber (MTCC 13366).
In summary, spores of A. niger melanoliber (MTCC 13366) secrete peptidomelanin during the germination process. Secretion is triggered after spores sense extracellular peptides via surface-associated peptide-spore interactions. We hypothesize that extracellular peptides present in the environment may function as nutritional cues, signaling substrate availability and thereby initiating the germination process, which includes the secretion of peptidomelanin. The preliminary work presented in this brief communication represents the first step in deciphering the peptidomelanin secretory pathway.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)