Keywords
Actinomycetes, Streptomyces, Abiotic stress, Salinity, Extreme pH, Thermal tolerance, Genomics, Biofertilizer
This article is included in the Bioinformatics gateway.
This article is included in the Genomics and Genetics gateway.
Actinomycetes, Streptomyces, Abiotic stress, Salinity, Extreme pH, Thermal tolerance, Genomics, Biofertilizer
Actinomycetes are free-living microorganisms, primarily soil saprophytes, dominated by Streptomyces. Their diversity and bioactivity depend on multiple environmental factors, and they fulfill key ecological functions such as nutrient cycling and availability, nitrogen fixation, siderophore production, and biocontrol of soil pathogens.1,2 Research on actinomycetes has shifted from focusing on their antibiotic production to recognizing their ecological and biotechnological potential. Current research delves into their metabolic versatility, genetic potential, and adaptability to extreme conditions.3 Actinomycetes are distributed in both common environments and extreme habitats (arid soils, caves, mountains, among others) thanks to their physiological and metabolic flexibility, including arthrospore formation and the production of secondary metabolites, which facilitates the discovery of new strains with bioactive properties.4 Therefore, natural elevations, due to their altitudinal gradients, thermal fluctuations and diverse soils, impose selective pressures (low temperatures, arid soils, high UV radiation and nutrient limitation) that favor the growth of actinomycetes with survival adaptations, so their isolations show high potential to discover new biotechnological alternatives.5
The coastal hills of northern Peru, such as the Cerro Campana hills (Peru), constitute a fog oasis highly dependent on winter drizzle, which favors the seasonal dynamics of vegetation on their sandy and sandy-clay soils. This creates a fragile ecosystem with marked altitudinal and microclimatic gradients that support a remarkable and locally relevant floristic biodiversity,6 but whose research and conservation efforts have been relatively isolated within the general landscape of studies on South American hills.7 Currently, there are no studies reporting the isolation of actinomycetes in the Cerro Campana hills; however, the presence of actinobacteria with high biotechnological potential has been demonstrated in arid soils. For example, Boukelloul et al.8 isolated 90 strains from rhizospheric and non-rhizospheric soils of arid Saharan regions, mainly from the genus Streptomyces, which showed antifungal activity against phytopathogens and plant growth-promoting activity. Similarly, Tan et al.9 describe the isolation of unique species of the genus Amycolatopsis from arid Australian soils, confirming that these environments constitute reservoirs of phylogenetically differentiated actinomycetes with potential for the production of secondary metabolites of biotechnological interest.
The study of adaptive mechanisms has been enhanced by the hybrid assembly of bacterial genomes, which has allowed for the acquisition of complete sequences. Functional annotation of these sequences using tools such as Prokka and NCBI-PGAP has facilitated the identification of genes associated with tolerance to abiotic stress (salinity, UV radiation, and extreme temperatures), revealing the genetic basis of adaptive mechanisms in bacteria from polyextreme environments.10 Furthermore, the integration of metagenomic assembly, binning, quality validation (CheckM), taxonomic classification (GTDB-Tk), and functional annotation allows for the reconstruction of genomes (MAGs) and the association of specific genes and metabolic pathways with ecological functions, such as carbon fixation, ammonium oxidation, or sulfur metabolism. This facilitates the identification of candidate genes involved in environmental adaptation and ecosystem resilience.11,12
In actinomycetes, key genes related to the biosynthesis of compatible solutes, such as ectA, ectB, and ectC, have been described. These genes are involved in ectoine production under osmotic stress,13 as well as genes associated with trehalose synthesis (otsA/otsB), Na+/H+ antiporters (nhaA, nhaP), and multication/proton complexes that maintain ionic homeostasis.14,15 Similarly, heat shock proteins (dnaK, groEL, groES), extracellular sigma factors (ESF), and universal stress proteins play a central role in regulating the cellular response to temperature and pH fluctuations.16,17 Identifying these genetic determinants is fundamental to understanding microbial resilience in extreme environments.
In this context, the genomic profiling of actinomycetes isolated from the Lomas Cerro Campana Conservation Area represents a relevant strategy for identifying genes associated with tolerance to salinity, temperature, and pH. The integration of genomic assembly, functional annotation, and comparative analysis allows not only the characterization of the genetic basis of abiotic adaptation but also the exploration of its potential application in agricultural and environmental biotechnology, particularly in the development of bio-inputs for degraded soils and production systems exposed to climate change.
Two rhizosphere soil samplings were carried out in the ACP–Lomas Cerro Campana, Trujillo, La Libertad, Peru. Climatic parameters were obtained from the POWER (Prediction Of Worldwide Energy Resources) v2.0.0 data access viewer. The first sample, corresponding to sampling “P” with two sampling points, P2 (7°59′31.454″ Latitude and 9°05′34.509″ Longitude) and P9 (7°59′10.997″ Latitude and 9°06′08.520″ Longitude), presented conditions of low light drizzle, with 83.31% relative humidity, 0.01 mm/day of precipitation and a UV index of 1.95. Meanwhile, the “Q” sampling with two sampling points Q3 (7°59′25.032″ Latitude and 79°05′21.215″ Longitude) and Q6 (7°59′23.358 Latitude y 79°05′25.076 Length) was carried out in warm and sunny conditions, with 84.56% relative humidity and a UV index of 2.80.
Rhizosphere soil samples were sieved using a 2-mm mesh. Subsequently, 10 g of soil were suspended in 90 mL of sterile 0.85% saline solution (SSE), homogenized manually, and serial decimal dilutions were prepared in SSE up to 10−7. The last three dilutions were surface-spread onto two selective media: Oat agar (ISP 3; HiMEDIA® M358)18 and Actinomycete Isolation Agar (AIA; Sigma aldrich, 17117-500G) supplemented with 0.1% nystatin (50 μg/mL; Sigma Aldrich, N1638-20ML). The plates were incubated at 30 °C for 7–15 days. Pure actinomycete cultures were stored in ISP 4 medium at 4 °C and in sterile 20% glycerol (Sigma aldrich, G9012-100ML) at −20 °C.19
For inoculant preparation, two colony-forming units (CFU) were selected from each actinomycete strain and incubated in actinomycete isolation broth at 30 °C, with shaking at 150 rpm, for 5 days. Subsequently, the bacterial biomass was recovered by centrifugation at 8000 rpm for 10 min, resuspended, and streaked onto nutrient agar plates, which were incubated at 30 °C for 48 h. Inoculum standardization was performed by counting (CFU/mL) and adjusting the suspension to a final concentration of 1 × 105 CFU/mL, which was used in subsequent assays.
Abiotic stress assessment: tolerance to salinity, temperature, and pH
The evaluation was performed using ISP3 solid medium as the basal substrate. Salinity tolerance was determined by supplementing the medium with NaCl (Sigma aldrich, S9888-500G) at concentrations of 3, 6, 9, 12, and 15% (w/v). Similarly, pH tolerance was evaluated in a range of 4 to 10. Temperature tolerance was determined by incubating the plates at 20, 30, 40, and 50 °C.20 Standardized inocula were streaked onto the medium adjusted to each abiotic condition. After 5 days of incubation, growth was assessed by counting (CFU/mL), emphasizing that the observed growth originated from the initial inoculum concentration.
Genomic DNA extraction was performed using the NucleoSpin™ Microbial DNA Mini Kit (Macherey-Nagel™, 740235.50), following the manufacturer’s instructions. From pure cultures of actively growing actinomycetes, 1–2 mL of cell biomass was collected and centrifuged at 10,000 × g for 5 min; the supernatant was discarded, and the pellet was washed with sterile PBS to remove any remaining medium. Subsequently, cell lysis was carried out by adding lysis buffer (MBL buffer), which contains detergent agents and chaotropic salts that promote membrane disruption. Additionally, a lysozyme pretreatment was included to optimize the disruption of the thick cell wall characteristic of Gram-positive actinomycetes, as previously reported to improve DNA yield in this bacterial group.21 Following lysis, binding buffer (MB Buffer) was added, and the mixture was transferred to silica columns, where the DNA was selectively adsorbed in the presence of high salt concentrations. The columns were subjected to successive washes with MW1 and MW2 buffers to remove proteins, polysaccharides, and other contaminants by centrifugation at 12000 × g. Finally, the DNA was eluted with 50–100 μL of low ionic strength ME Buffer. The concentration and purity of the extracted DNA were determined by spectrophotometry (A260/A280 ratio), and its integrity was verified by agarose gel electrophoresis.22
Sequencing was performed using the MinION platform (Oxford Nanopore Technologies, ONT), employing the Rapid Barcoding Kit 24 V14 (SQK-RBK114.24), adapting the workflow described for the RBK110 kit to our experimental conditions.23 For each sample, the genomic DNA was not diluted due to its low initial concentration. Each isolate was labeled with a distinct barcode, allowing multiplexing in a single flow cell. The barcode tagging and incorporation reaction was performed according to the manufacturer’s instructions, with controlled incubations for enzymatic fragmentation and adapter binding. Subsequently, the barcode libraries were pooled and prepared for loading by adding the rapid adapter (RAP) and the corresponding sequencing reagents. A new R9.4.1 flow cell was used, which had more than 800 active pores at the time of the pre-run check. The loading was performed via the SpotON port, avoiding the introduction of bubbles to preserve the integrity of the nanopore matrix, in accordance with the technical recommendations of the original protocol. The sequencing run was executed using MinKNOW software with high-precision base calling enabled and automatic barcode demultiplexing. The generated reads were filtered by minimum quality (Qscore ≥7) and minimum read length ≥ 1 kb.
The bacterial genomes were assembled from long-read sequencing data using Flye v2.9.5, a repetitive graph-based assembler designed specifically for third-generation data (Nanopore/PacBio).24 Assembly was performed using parameters optimized for prokaryotic genomes and automatic estimation of genome size, a procedure recommended to maximize contig continuity in Actinobacteria genomes, which typically range in size from 6 to 10 Mb. Assembly quality was evaluated based on N50, contig count, and G + C content using QUAST (v5.2.0).25 The assembled genomes were then annotated structurally and functionally using Prokka v1.13.4, an automated tool for rapid annotation of prokaryotic genomes that integrates ORF prediction with Prodigal, identification of rRNA and tRNA genes, and functional assignment based on databases such as UniProt and RefSeq.26
The genome sequence data for the four Streptomyces spp. strains isolated from the Lomas del Cerro Campana have been deposited in DDBJ/ENA/GenBank under the accession numbers JBVQPS000000000 (P9.2), JBVQPT000000000 (P2.1), JBVQPU000000000 (Q3.3), and JBVQPV000000000 (Q6.2). All sequences are linked to BioProject PRJNA1430095 and to BioSample accession numbers SAMN56266636, SAMN56300581, SAMN56300630, and SAMN56300674, respectively, at the National Center for Biotechnology Information (NCBI).
The genome sequence data were uploaded and taxonomically analyzed using digital DNA–DNA hybridization (dDDH) through TYGS (Type Genome Server).27 Genome comparisons were carried out with GBDP (Genome Blast Distance Phylogeny).28 In addition, dDDH values and their confidence intervals were estimated using GGDC v3.0 (Genome-to-Genome Distance Calculator).28,29 The phylogenetic tree was then generated with FASTME v2.1.6.1 based on 100 pseudo-bootstrap replicates,30 rooted at the midpoint, and visualized using PhyD3.31
Functional annotation and category assignment for Clusters of Orthologs (COGs) were performed using eggNOG-mapper v2,32 based on the eggNOG v5.0 orthology database.33 They were grouped into four functional categories for analysis: (i) thermal adaptation and proteostasis, (ii) pH homeostasis and ion flux, (iii) osmotic tolerance and structural integrity, and (iv) regulation and environmental sensing. Based on this annotation, genetic determinants linked to tolerance to pH, salinity, and temperature were identified; the selection criteria were based on identifying determinants that demonstrated exclusivity in certain isolates or functional complementarity among them.
To determine the functional proximity of these selected genes, heatmaps were generated using the Heatmap2 tool.34 This analysis made it possible to identify patterns of co-occurrence and divergence among the isolates by applying a dual hierarchical clustering method based on Euclidean distance and the complete linkage method.
Four pure cultures of actinomycetes were isolated, in which in sampling P, 2 isolates were obtained, named P2.1 and P9.2, while in sampling Q the isolates Q3.3 and Q6.2 were obtained ( Figure 1). In Figure 1, the four strains exhibit macromorphology consistent with Streptomyces, as they display dry/compact colonies, radial growth, differentiation between substrate mycelium and aerial mycelium, and variation in pigmentation and the degree of sporulation.
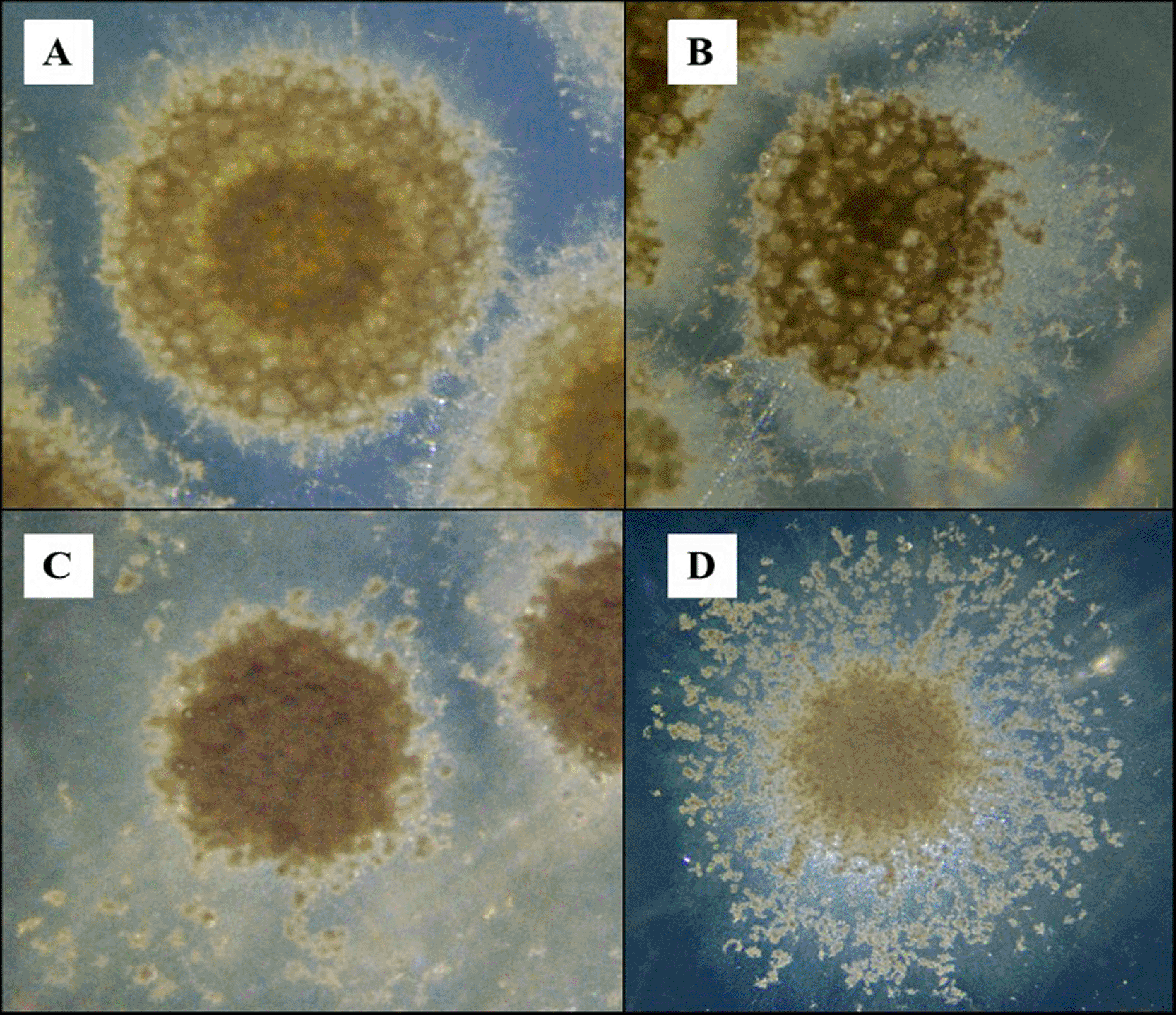
Stereomicroscopic view of colonies showing various growth patterns, including aerial mycelium formation, spore pigmentation, and margins of (A) Q3.3, (B) P2.1, (C) Q6.2, and (D) P9.2. The strains were inoculated onto actinomycete isolation agar and incubated at 30 °C for 7 days.
Figure 2 shows that the growth of strains Q3.3, P2.1, Q6.2, and P9.2 varied depending on temperature, pH, and NaCl concentration. With regard to temperature (Figure 2A), all strains grew between 20° and 40 °C, although at different rates. In general, the highest growth was observed at 30 °C, especially in P9.2, which reached the highest CFU/mL values, followed by Q3.3. Significant growth was also recorded at 20 °C, mainly in Q6.2 and P9.2. In contrast, growth decreased at 40 °C, and at 50 °C it was very low or nearly absent, although Q3.3 retained some tolerance. With regard to pH (Figure 2B), all four strains were able to grow over a wide range, from pH 4 to 10, indicating tolerance to acidic, neutral, and alkaline conditions. However, optimal growth was observed mainly between pH 6 and 8. Strain P9.2 exhibited the highest count across nearly the entire evaluated range, particularly at pH 7 and 8. Q6.2 also showed good growth, especially at values close to neutrality and slight alkalinity. Meanwhile, Q3.3 and P2.1 grew across the entire range, although with lower counts at the pH extremes.
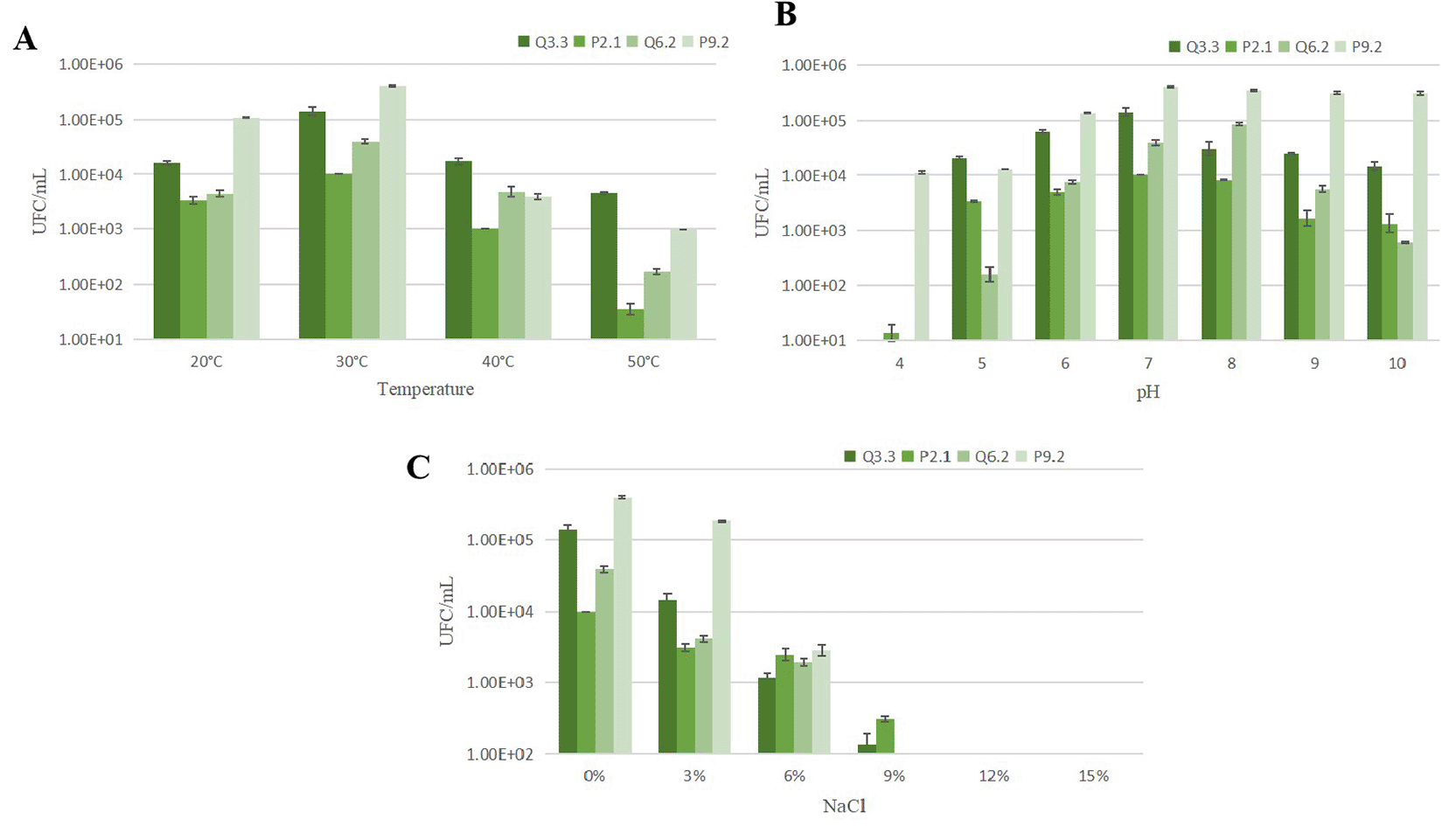
With regard to salinity (Figure 2C), growth was highest in the absence of NaCl or at low concentrations. The highest values were recorded at 0% NaCl, again with a predominance of P9.2 and Q3.3. As the salt concentration increased to 3% and 6%, growth decreased progressively in all isolates, although growth was still observed. At 9% NaCl, growth was low, and only a few isolates, such as P2.1 and, to a lesser extent, Q3.3, maintained detectable counts. At 12% and 15% NaCl, no growth was observed, indicating limited tolerance to high salt concentrations.
These results indicate that P9.2 was the isolate with the best overall performance under varying temperature and pH conditions, while all isolates showed a preference for moderate temperatures (30 °C), near-neutral pH, and low salinity.
Among the molecular characteristics of the genomes of the four strains listed in Table 1, the genome sizes of strains P9.2 and P2.1 range from 7.3 Mbp to 8.4 Mbp, respectively. The G + C content showed high consistency across the strains, with Q3.3 and P9.2 having the highest percentages. Regarding the continuity and quality of the assembly, two strains exhibit a single contig of 6.8 Mbp and 7.8 Mbp, in contrast to the other two, which consist of fragmented structures of 30 and 120 contigs, with varying assembly quality values; the N50 of the single-contig genomes is greater than 6. 8 Mbp, while the fragmented genomes have values of 150,755 bp and 76,822 bp. Completeness levels exceed 98.1%, with contamination values below 1.35% in strains Q3.3, P9.2, and P2.1; in contrast, strain Q6.2 exhibits up to 4.68% contamination. The number of coding sequences is proportional to genome size, with 7,335 CDS identified in P2.1, which has a larger genome. Regarding transcription and translation RNAs, they exhibit a total RNA (tRNA and rRNA) count of between 91 and 107 molecules per genome.
The phylogenetic analysis based on whole genomes was performed using the Type (Strain) Genome Server (TYGS), revealing taxonomic divergence among the strains and grouping them into two clades within the genus Streptomyces.
Strains Q6.2 and P9.2 have 100% node support, forming a monophyletic group ( Figure 3), and are positioned close to Streptomyces lusitanus JCM 4785. Despite their proximity on the tree, analysis of the digital DNA-DNA hybridization (dDDH) values and G + C content of strains Q6.2 and P9.2 showed dDDH values of 41.2% and 41.7% and G + C contents of 0.11 and 0.13, and the closest type strains were S. albogriseolus JCM 4616 and S. viridodiastaticus JCM 4536, respectively. Strains Q3.3 and P2.1 were grouped into a second independent clade, close to Streptomyces plicatus JCM 4504 and Streptomyces vinaceusdrappus JCM 4529, with support of 66% and 100%, respectively; furthermore, strain Q3. 3 was identified as the type strain closest to S. geysiriensis JCM 4962 with 87.7% dDDH and 0.24% G + C, and strain P2.1 as closest to S. vinaceusdrappus JCM 4529 with 86.8% dDDH and 0.12% G + C.
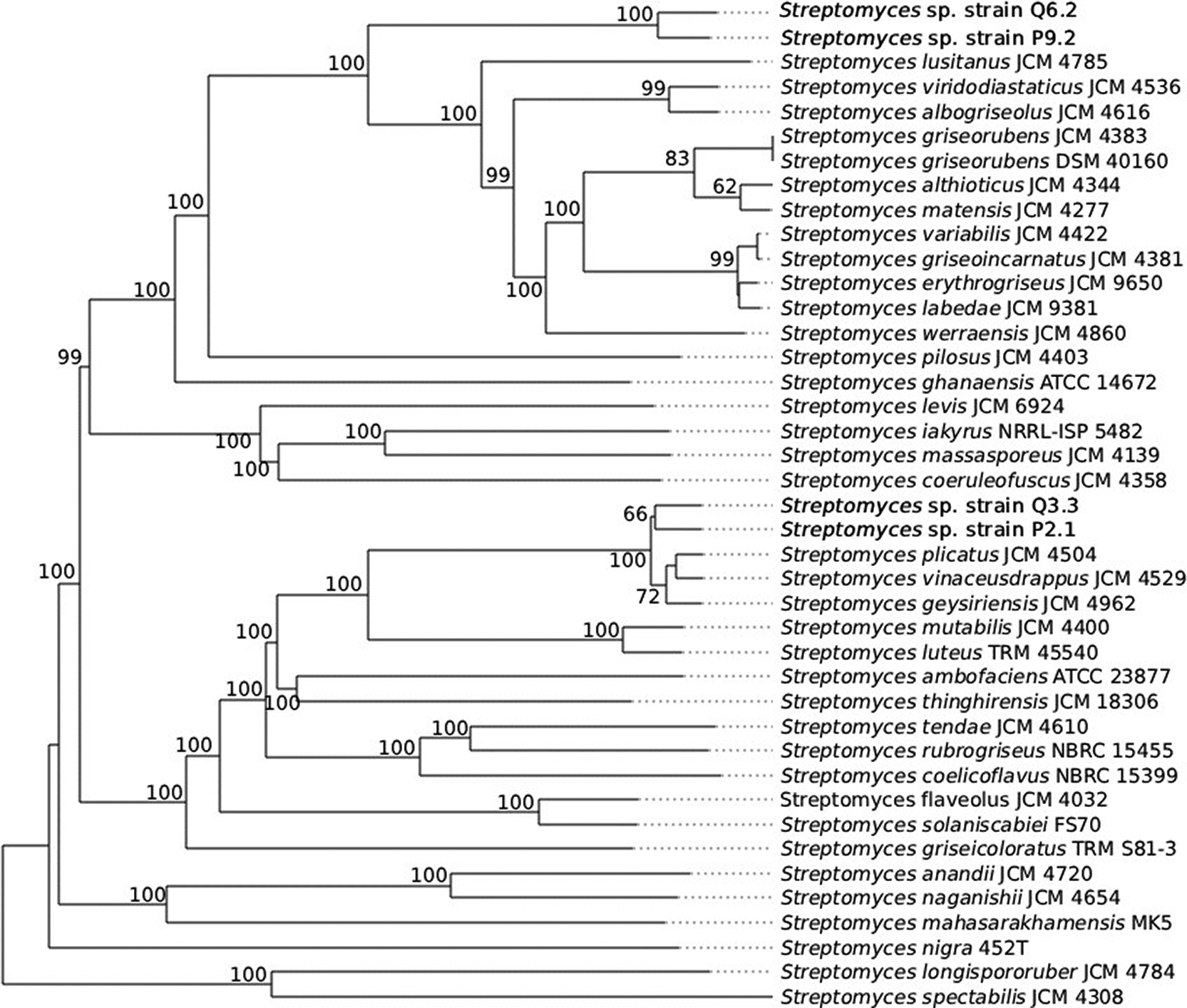
The numbers on the nodes represent bootstrap support values based on genome-wide comparisons. Streptomyces spectabilis JCM 4308 was used as an outgroup.
The functional classification of Orthologous Groups (COGs) reveals genes associated with environmental adaptation responses, including those related to temperature, pH, and salinity stress. In thermal adaptation and proteostasis ( Figure 4A), genes related to Replication, recombination, and repair (L) were abundant, including strain P2.1 with 292 genes; in Translation and ribosomal structure (J), similar values were observed, ranging from 222 (P9.2) and 236 (Q6.2) genes; regarding post-translational modification and chaperones (O), between 160 and 169 genes were found in strains P2.1, Q6.2, and Q3.3, while 150 genes were found in P9.2. In response to a state of pH and ion flux homeostasis ( Figure 4B), the most prominent function is Energy production and conversion (C), with strain Q6.2 having the highest number of genes (341), followed by strain P2.1 with 337 genes; In the transport and metabolism of inorganic ions (Q), there was greater variability in the number of genes, ranging from 188 (P9.2) to 235 (P2.1); defense mechanisms (V) showed similar ranges, from 146 (Q6.2) to 168 (P2.1) genes. In response to osmotic stress and envelope integrity ( Figure 4C), the highest number of genes was found in Carbohydrate transport and metabolism (G) with 466 (P2.1) and 460 (Q3.3) genes, and amino acid transport and metabolism (E) with 413 (P9.2) and 447 (P2.1) genes; the number of genes for cell wall and membrane biogenesis (M) was consistent between 257 (P9.2) and 271 (Q6.2). The functional category of Transcription (K) within Environmental Perception and Regulation ( Figure 4D) had the highest number of genes in the entire analysis, with values of 755 (P2.1), 733 (Q3.3), 712 (Q6.2), and 672 (P9.2); in Signal Transduction (T), the number of genes ranged from 323 (P9.2) to 360 (P2.1); genes linked to Secondary Metabolism (Q) showed similar values among the strains P2.1 (187), Q6.2 (181), and Q3.3 (187), with the P9.2 strain differing at 160 genes.
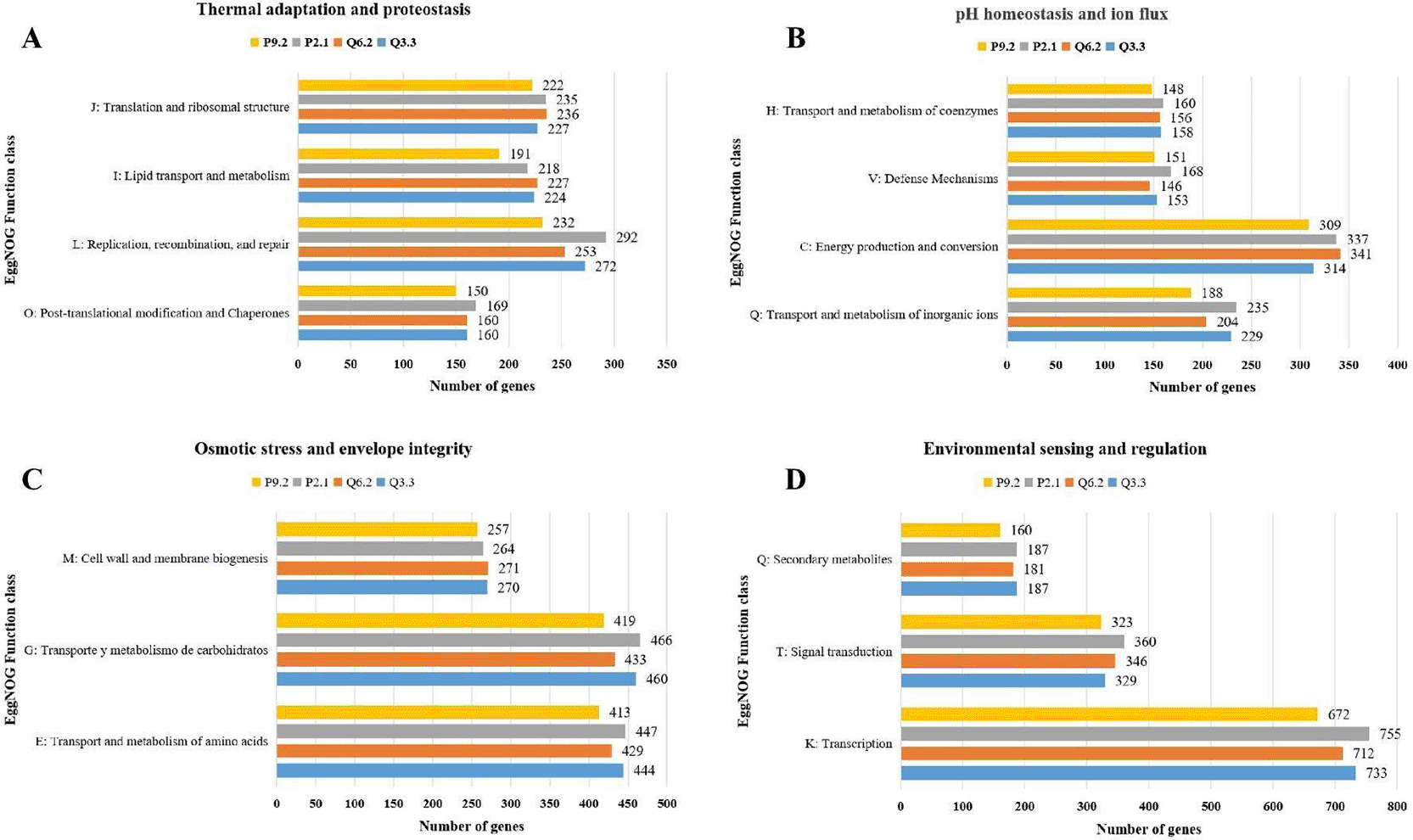
The categorization is based on the Clusters of Orthologs (COGs) obtained using eggNOG-mapper. (A) Thermal adaptation and proteostasis: Chaperone systems, DNA repair, and membrane fluidity modification. (B) pH homeostasis and ion flux: Bioenergetic machinery and inorganic ion transport systems for acid-base balance. (C) Osmotic stress and envelope integrity: Biosynthetic pathways for compatible solutes and cell wall biogenesis. (D) Environmental sensing and regulation: Signal transduction networks and transcriptional factors that coordinate the adaptive response.
Among the genes involved in heat shock response and proteostasis ( Figure 5A), the gyrA, groL2, ssb, and dnaK genes are conserved across all four strains; the polA gene is most abundant across all strains; the hup gene is most highly expressed in strain P2.1; among the specialized chaperones, the clpB and hsp18 genes are upregulated in strains Q6.2 and P9.2, while their expression is reduced in the other strains. Compared to the state of pH homeostasis ( Figure 5B), the katG, furA, and dps genes are clustered together and present in P2.1 and Q3.3; the ctaD gene is found only in strain Q6.2; similarly, the mgtE gene is specific to Q3.3; the atpA gene is present in all strains. The genes involved in osmotic adaptation ( Figure 5C), such as betT, proC, glnQ, algD, and glgB, are grouped together as a profile unique to strain P9.2; the ectB gene is present only in strain Q6.2, while the mscS, otsB1, and cfa genes are grouped together and present in strain P2.1.
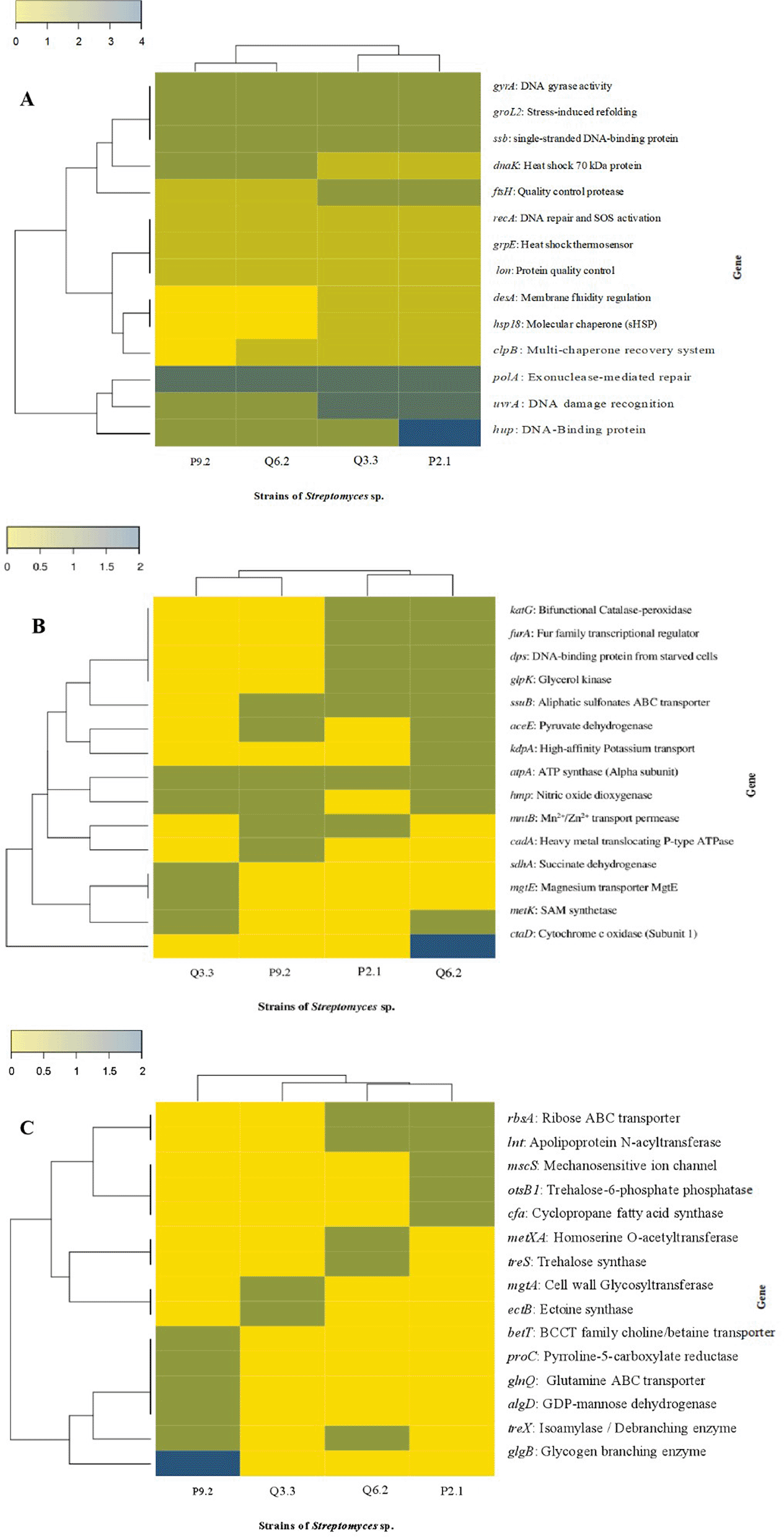
The distribution of key genes for tolerance to: (A) heat shock and proteostasis, (B) pH homeostasis, and (C) osmotic adaptation is shown. The dendrograms indicate the functional proximity among the strains (Q3.3, Q6.2, P2.1, and P9.2) based on the Euclidean distance metric. Color intensity represents gene abundance normalized by row scaling.
The morphological characteristics exhibited by the strains are typical forms that develop in culture media such as AIA.35 Strain Q3.3 formed a circular colony with a yellowish-brown center, a rough surface, and abundant peripheral aerial mycelium with a cottony-granular appearance. P2.1 exhibited a more compact, dark brown colony with a nodular surface and an irregular margin. Q6.2 showed a dense, matte colony with a uniform brown hue, sparse aerial mycelium, and a smooth to slightly wavy margin. In contrast, P9.2 exhibited the greatest development of aerial mycelium, with radial growth, a star-like appearance, and a filamentous margin. Taken together, these features are consistent with the typical macromorphology described for isolates of the genus Streptomyces on solid medium.36
In Figure 2, the highest overall growth was observed between 30 °C and a pH range of 6–8, while growth decreased markedly at 50 °C and at high NaCl concentrations. This pattern is consistent with reports for various Streptomyces strains, whose optimal growth ranges are typically between 20–40 °C, with little or no growth at 50 °C.37 Regarding temperature, the data suggest that the four strains are predominantly mesophilic, with an optimum near 30 °C, especially in P9.2 and Q3.3. This response is consistent with other studies in which Streptomyces showed best growth around 28–30 °C, while higher temperatures caused a significant reduction in biomass or metabolic activity; for example, Lertcanawanichakul and Sahabuddeen38 reported that Streptomyces sp. KB1 grew between 25 and 37 °C, with an optimum at 30 °C; similarly, strain PBR11 exhibited maximum growth at 28 °C.39 Therefore, the decline observed at 40 °C and, above all, at 50 °C, indicates that these strains are not thermophiles, but rather microorganisms adapted to moderate environmental conditions.
The genomes of the four Streptomyces strains isolated from the Lomas del Cerro Campana ACP are of high quality ( Table 1), with assembly completeness exceeding 98% and contamination levels below 5%. Furthermore, the approximate genomic size of 8 Mbp and G + C content exceeding 72% are characteristics typical of the genus Streptomyces, reflecting their vast metabolic and adaptive capacity in complex environments.40 Additionally, the high number of tRNA and rRNA genes are genomic features that correlate with a rapid growth response following long periods of dormancy; in arid environments, water availability is limited, and thus the presence of robust protein complexes allows these Streptomyces to reactivate their metabolism upon contact with water, giving them a competitive advantage over other less adaptable taxa.41,42
Strains P2.1 and Q3.3 form a monophyletic clade ( Figure 3) with S. vinaceusdrappus JCM 4529 and S. geysiriensis JCM 4962, with dDDH values of 86.8% and 87.7%, respectively, confirming their affiliation with the Streptomyces species, as they exceed the 70% species-delimiting threshold.43 In contrast, strains P9.2 and Q6.2 are most closely related to S. viridodiastaticus JCM 4536 and S. albogriseolus JCM 4616, respectively, whose maximum dDDH value is 41.7%, which is well below the species threshold. This, combined with the phylogenetic tree topology—where the strains are located on an external branch with 100% node support—suggests that these strains represent new species. These potential new Streptomyces species are likely due to the geographic distribution and environmental conditions of the Lomas del Cerro Campana ACP, where mutations may have developed that drive these genomic changes.44
Functional classification using eggNOG-mapper reveals that the Co-Orthologous Groups (COGs) are linked to different metabolic pathways and the number of genes associated with abiotic stress tolerance in Streptomyces, enabling them to colonize microbiomes with fluctuating nutrient availability.45 In terms of thermal adaptation and proteostasis ( Figure 4A), the P2.1 strain stands out for having 292 genes involved in replication and repair; combined with its larger genome size ( Table 1), these genes likely act as resilience mechanisms against damage caused by UV radiation and temperature fluctuations, such as those that occur in arid soils.46 Furthermore, they not only help mitigate heat stress but also maintain enzymatic function during periods of desiccation and rehydration.47 In response to pH and ion flux stress ( Figure 4B), there is a notable abundance of genes involved in energy production, due to the high demand for ATP required to maintain active electrochemical gradients under optimal conditions.48
In response to osmotic stress and membrane integrity ( Figure 4C), the largest number of genes are associated with the transport and metabolism of carbohydrates, which could include trehalose and ectoine as an initial osmoregulatory mechanism; meanwhile, the transport of amino acids such as proline would act as a multifunctional cytoprotective agent.49 The cell wall’s biosynthetic capacity ensures that it maintains its structural integrity in the face of sudden changes caused by external factors.50 The environmental and regulatory sensing pathway ( Figure 4D) comprises the most densely packed region of the genome, which is regulated by RNA polymerase sigma factors—a characteristic of Streptomyces that enables them to detect even minimal stimuli and trigger adaptive responses, such as the activation of secondary metabolites under stress.51
The four strains share the core of the folding systems that enable them to maintain protein functionality in the face of heat shock and proteostasis ( Figure 5A), due to the presence of dnaK (Hso70), dnaJ (Hsp40), and groEL/groES (Hsp60/Hsp10), which identify and isolate proteins to prevent interference during folding.52,53 In addition, the grpE gene enables DnaK to release folded proteins and restart the catalytic cycle, thereby maintaining an active and continuous response to this stress.54 In addition, the ftsH gene ensures the quality of integral membrane proteins, thereby ensuring ion transport.55 The Q3.3, Q6.2, and P2.1 strains carry clpB (Hsp100), a deaggregase that allows proteins affected by heat shock to be reintegrated into the folding cycle; exclusively, strain Q3.3 carries the htpG gene (Hsp90), which acts as a chaperone to maintain the stability of signaling proteins and receptors during prolonged stress.56 In strains Q6.2 and P2.1, the clp and lon genes function by degrading proteins irreversibly damaged by heat stress, a strategy for protein quality control and the avoidance of cytotoxic aggregates.41 The P2.1 and Q3.3 strains possess a fatty acid desaturase, desA, which maintains the membrane’s homeoviscosity by adjusting the lipid composition to keep the fluidity constant, allowing the cell to adapt to rapid temperature changes.57
In pH homeostasis ( Figure 5B), the kdpA gene, which is unique to strain Q6.2, responds to the recovery of cell turgor following an osmotic shift by allowing potassium to enter the cell to stabilize the cytoplasm.58 In addition, the ctaD gene responds by generating proton gradients to maintain active transport under stressful conditions.59 In addition, the mgtE gene plays a role in the transport of specific divalent cations such as magnesium, helping to maintain the integrity of ribosomes and the stability of cellular metabolism, and acts in conjunction with the metK gene, which modifies membrane lipids through methylation to reduce passive proton permeability.60 In the P9.2 strain, the cadA gene enables it to act as a heavy metal detoxification system under acidic conditions,53 along with the mntB gene, which enables it to uptake cations such as magnesium and zinc that are typically unavailable at alkaline pH levels.61 Furthermore, the ssuB gene, which it shares with strains Q6.2 and P2.1, enables it to acquire sulfur—a vital component for amino acid synthesis in response to acid-base stress.62 Strains Q3.3, P9.2, and Q6.2 share the hmp gene, which is responsible for mitigating nitrosative stress caused by pH fluctuations; the aceE gene prevents excessive internal acidification by producing organic acids through pyruvate metabolism.63
The greatest genetic divergence was observed in osmotic adaptation ( Figure 5C); strain Q3.3 possesses the ectB gene, which is responsible for the biosynthesis of ectoine—a compound that acts as a stabilizer for protein hydration under water stress—and, together with the mgtA gene, stabilizes cell wall glycosylation in the face of ionic toxicity.60,64 The P9.2 strain, which carries the proC and betT genes, synthesizes L-proline as an osmoprotectant and captures glycine betaine, respectively, enabling it to withstand salt shocks effectively and efficiently.65 The glgB gene enables glycogen storage, which, together with the energy transporter encoded by the glnQ gene, facilitates the active transport of nutrients during osmotic stress.66,67 In addition, the algD gene is responsible for the secretion of alginate, which forms a matrix that retains water and protects the colonies from drying out.68 The P2.1 strain, which carries the otsB1 gene, produces trehalose, which prevents protein denaturation during extreme desiccation; furthermore, the mscS and cfa genes act as a mechanosensitive channel, preventing cell lysis due to osmotic shock, and encode membrane phospholipids, thereby reducing sodium ion permeability.69,70
Four Streptomyces strains were identified that possess a genetic repertoire linked to mechanisms for adapting to extreme abiotic conditions, notably genes related to replication, recombination, and repair, as well as chaperones and post-translational modifications involved in the heat response; furthermore, it was demonstrated that their adaptations to arid environments do not depend on a single gene, but rather on a set of complementary mechanisms. In response to heat shock and proteostasis, conserved genes such as gyrA, groL2, ssb, dnaK, and polA—associated with DNA stability, protein folding, and cellular repair—stood out; furthermore, clpB and hsp18 were prominent in Q6.2 and P9.2, while hup showed greater intensity in P2.1. The genes katG, furA, dps, atpA, ctaD, and mgtE were identified as being involved in pH homeostasis and ion flux; these are associated with antioxidant defense, iron regulation, energy balance, and cation transport, ctaD stood out in Q6.2 and mgtE in Q3.3. In osmotic adaptation, the following genes were identified betT, proC, glnQ, algD, glgB, ectB, mscS, otsB1, and cfa, which are associated with the biosynthesis of osmoprotectants, the uptake of compatible solutes, energy reserves, protection against desiccation, and membrane stability. The main finding was that P9.2 exhibited the best overall phenotypic performance under varying temperature and pH conditions and, furthermore, displayed a unique osmotic profile involving betT, proC, glnQ, algD, and glgB; while Q6.2 and P9.2 might even represent new species within the genus Streptomyces, reinforcing their potential biotechnological value for bio-inputs applicable in degraded soils and climate change scenarios.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)