Keywords
Caryophyllaceae, chloroplast genome, endemic species, Dianthus
This article is included in the Genomics and Genetics gateway.
Caryophyllaceae, chloroplast genome, endemic species, Dianthus
This revised version includes several updates to improve methodological clarity, figure presentation, and overall readability. In the Methods section, we added a clearer description of how clean reads were obtained from raw sequencing data before plastome assembly with NOVOPlasty. We also specified that nucleotide diversity was calculated using aligned plastome sequences from nine Dianthus species, including D. helenae. Figure 1 was revised by adding panel labels and subtitles to clarify the habit, habitat, and floral details of D. helenae. The chloroplast genome map was also replaced with the original uncropped version, and its caption was expanded to explain the LSC, SSC, IRa, and IRb regions, gene transcription directions, functional gene colors, GC/AT content, region boundaries, and GC/AT skew curves. Minor formatting and wording corrections were made throughout the manuscript, including the consistent italicization of scientific names. These revisions improve the transparency and presentation of the article, while the main results and conclusions remain unchanged.
See the authors' detailed response to the review by Hoang Dang Khoa Do
Dianthus L. is one of the major genera of Caryophyllaceae and represents a taxonomically complex and evolutionarily important lineage within Caryophyllales. Recent taxonomic work has provided the first broad phylogenetic framework for the genus using plastid markers together with nuclear ITS data and has recognized 384 accepted species, confirming that Dianthus is far more diverse than traditionally assumed (Fassou et al., 2022). At the same time, phylogenetic reconstruction in the genus remains challenging. Previous studies have shown that genetic distances among many Dianthus taxa are very low, species-level relationships are often weakly resolved, and plastid haplotypes may be deeply shared among related species, suggesting rapid radiation and a complex evolutionary history (Fassou et al., 2022). More recent molecular analyses have nevertheless begun to recover major lineages within the genus. Combined plastid and nuclear datasets support the monophyly of Dianthus and indicate clear geographic and sectional structuring, while plastome-based analyses have resolved two strongly supported major clades among sampled species, demonstrating that chloroplast data are informative for understanding evolutionary relationships in the genus (Lin et al., 2022; Mnxati & Mankga, 2025).
Beyond its phylogenetic importance, Dianthus is also notable for its ornamental, economic, and medicinal value. Several species are widely cultivated, and a number of taxa have long been used in traditional medicine, making the genus relevant not only to systematics but also to applied plant science (Lin et al., 2022; Raman & Park, 2015; Meng et al., 2023). However, morphological similarity among species, frequent hybridization, and the limited availability of genomic resources have complicated species delimitation and the interpretation of evolutionary relationships in Dianthus (Lin et al., 2022; Meng et al., 2023). In this context, complete chloroplast genomes provide a particularly useful source of evidence because they are generally conserved in structure and gene content, maternally inherited in most angiosperms, and widely used in comparative genomics, DNA marker development, and phylogenetic reconstruction (Nikitina et al., 2025; Alieva et al., 2025; Tojiboeva et al., 2025; Dekhkonov et al., 2025; Ergashov et al., 2026a, b, c). Previous plastome studies in Dianthus have shown that chloroplast genomes are structurally conserved and phylogenetically informative, but taxon sampling remains limited and is still insufficient to fully clarify relationships across the genus (Lin et al., 2022; Yang et al., 2021). Dianthus helenae Vved., an accepted species originally described in 1941, remains poorly studied at the genomic level (Fassou et al., 2022). Therefore, characterization of its complete chloroplast genome will not only provide a new genomic resource for the genus, but will also help clarify its phylogenetic placement within Dianthus, contribute to comparative plastome studies, and support future taxonomic and conservation research on this endemic medicinal species from the Nuratau Mountains of Uzbekistan.
Fresh leaves of Dianthus helenae Vved. were collected from a wild population in the Nuratau Range, Uzbekistan ( Figure 1). Species identification was carried out by N. Beshko, and the voucher specimen is deposited in the National Herbarium of Uzbekistan (TASH) under accession number TASH139868.

A – Close-up of reproductive shoots of D. helenae; B – Whole plant in its natural habitat; C – Close-up of flowering individuals. Various individuals of D. helenae in a shrub–grassland-steppe community on rocky and fine-soiled slopes in the Ustuksay area, Nuratau Range, Uzbekistan (approximately 1450 m a.s.l.; 4 July 2024; photograph by N. Beshko).
Total genomic DNA was extracted from approximately 100 mg of fresh leaf tissue using a Tiangen Plant Genomic DNA Kit (Tiangen Biotech Co., Beijing, China; Cat. No. DP305) following the manufacturer’s protocol. Sequencing libraries were prepared using ~1 μg of purified DNA with the NEBNext Ultra II DNA Library Prep Kit for Illumina (New England Biolabs, USA; Cat. No. E7645S), including fragmentation to ~350 bp, end repair, adapter ligation, and PCR amplification. Library quality and fragment size distribution were assessed using an Agilent 5400 system. Qualified libraries were sequenced on an Illumina platform at Novogene Bioinformatics Technology Co.
Raw reads were first quality-filtered to remove adapter sequences, low-quality reads, and reads containing ambiguous bases. The resulting clean reads were then used for de novo plastome assembly with NOVOPlasty (Dierckxsens et al., 2017). Gene annotation was performed in Geneious Prime using Dianthus chinensis (GenBank accession OP136025) as the reference genome and manually verified for start and stop codons as well as exon–intron boundaries (Kearse et al., 2012). A circular chloroplast genome map was generated using OGDRAW (Greiner et al., 2019). The assembled chloroplast genome was deposited in GenBank under accession number PZ251004.
Nucleotide diversity (Pi) was calculated using DnaSP v6.12.03 based on the aligned plastome sequences of eight Dianthus species, including D. helenae (Rozas et al., 2017). A sliding-window analysis was performed with a window length of 1000 bp and a step size of 500 bp.
For phylogenetic reconstruction, 22 plastome sequences representing nine Dianthus species and one outgroup taxon (Petrorhagia saxifraga) were retrieved from NCBI GenBank ( Table 1). Complete chloroplast genomes were aligned using MAFFT (Katoh & Standley, 2013). Maximum likelihood analysis was performed in RAxML with 1,000 bootstrap replicates (Stamatakis, 2014). The GTR + G model was selected using jModelTest v2.1.4 under the Akaike Information Criterion (Darriba et al., 2012).
Bold accession means sequenced sample for this study.
The chloroplast genome map of Dianthus helenae revealed a typical circular quadripartite structure, consisting of a large single-copy (LSC) region of 82,836 bp, a small single-copy (SSC) region of 17,105 bp, and a pair of inverted repeats (IRa and IRb) of 24,884 bp each, giving a total genome length of 149,709 bp ( Figure 2). The genome contained the usual set of plastid genes involved in photosynthesis, ATP synthesis, transcription, and translation, including protein-coding genes, transfer RNAs, and ribosomal RNAs. As shown in the map, genes were distributed on both strands, while several genes located in the IR regions were duplicated, including rRNA and some tRNA/protein-coding genes. This agrees well with comparative studies showing that Dianthus chloroplast genomes are highly uniform in overall architecture, generally falling within a narrow size range and containing largely conserved gene sets, with only limited variation at IR/SC boundaries (Lin et al., 2022; Meng et al., 2023; Wicke et al., 2011; Daniell et al., 2016; Yang et al., 2021).
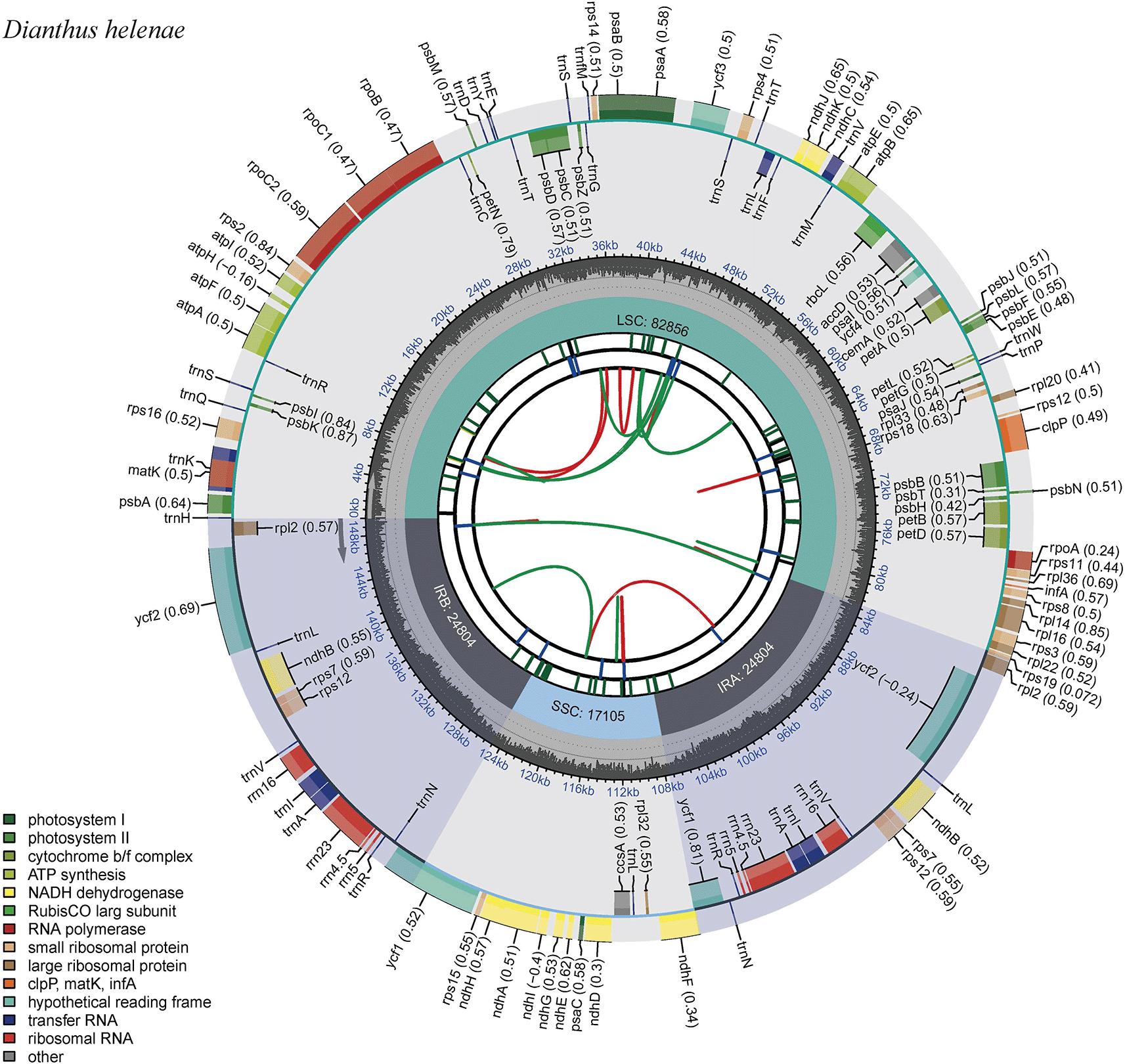
The map shows the organization of genes in the large single-copy region (LSC), small single-copy region (SSC), and two inverted repeat regions (IRa and IRb). Genes located outside the outer circle are transcribed clockwise, whereas genes located inside the circle are transcribed counterclockwise. Genes are color-coded according to their functional categories. The inner gray ring represents nucleotide composition, with darker gray indicating GC content and lighter gray indicating AT content. The green boundary lines indicate the junctions between the LSC, SSC, IRa, and IRb regions. The red and green curves in the innermost circle show GC skew and AT skew, respectively.
Phylogenetic analysis based on complete chloroplast genome sequences strongly supports the placement of D. helenae within the genus Dianthus ( Figure 3). The maximum likelihood tree reveals that D. helenae forms a well-supported clade closely related to D. chinensis and D. caryophyllus, with high bootstrap values (≥100), indicating robust phylogenetic inference.

Phylogenetic relationships inferred from complete chloroplast genome sequences. Bootstrap values are shown at the nodes. Petrorhagia saxifraga was used as the outgroup.
The clustering pattern is consistent with previous plastome-based phylogenies, confirming the reliability of whole chloroplast genome data fogeir resolving relationships within Caryophyllaceae. The distinct positioning of D. helenae suggests its independent evolutionary trajectory within the genus. This is broadly compatible with previous whole-plastome analyses, which also resolved two major, well-supported chloroplast groups within Dianthus, one centered on D. caryophyllus, D. barbatus, and D. gratianopolitanus, and the other on D. superbus, D. chinensis, and D. longicalyx (Lin et al., 2022).
Sliding window analysis of nucleotide diversity (Pi) revealed heterogeneous variation across the plastome ( Figure 4). The SSC region exhibited higher variability compared to LSC and IR regions, consistent with patterns observed in other angiosperms. Several highly variable regions were identified, including coding regions such as ycf1, rpl32 and intergenic spacers ndh-ndhE, rps16-trnQ-UUG. These hotspots represent potential candidate regions for molecular marker development and phylogeographic studies. In contrast, IR regions showed significantly lower nucleotide diversity, reflecting their conserved nature and the stabilizing effect of gene duplication.
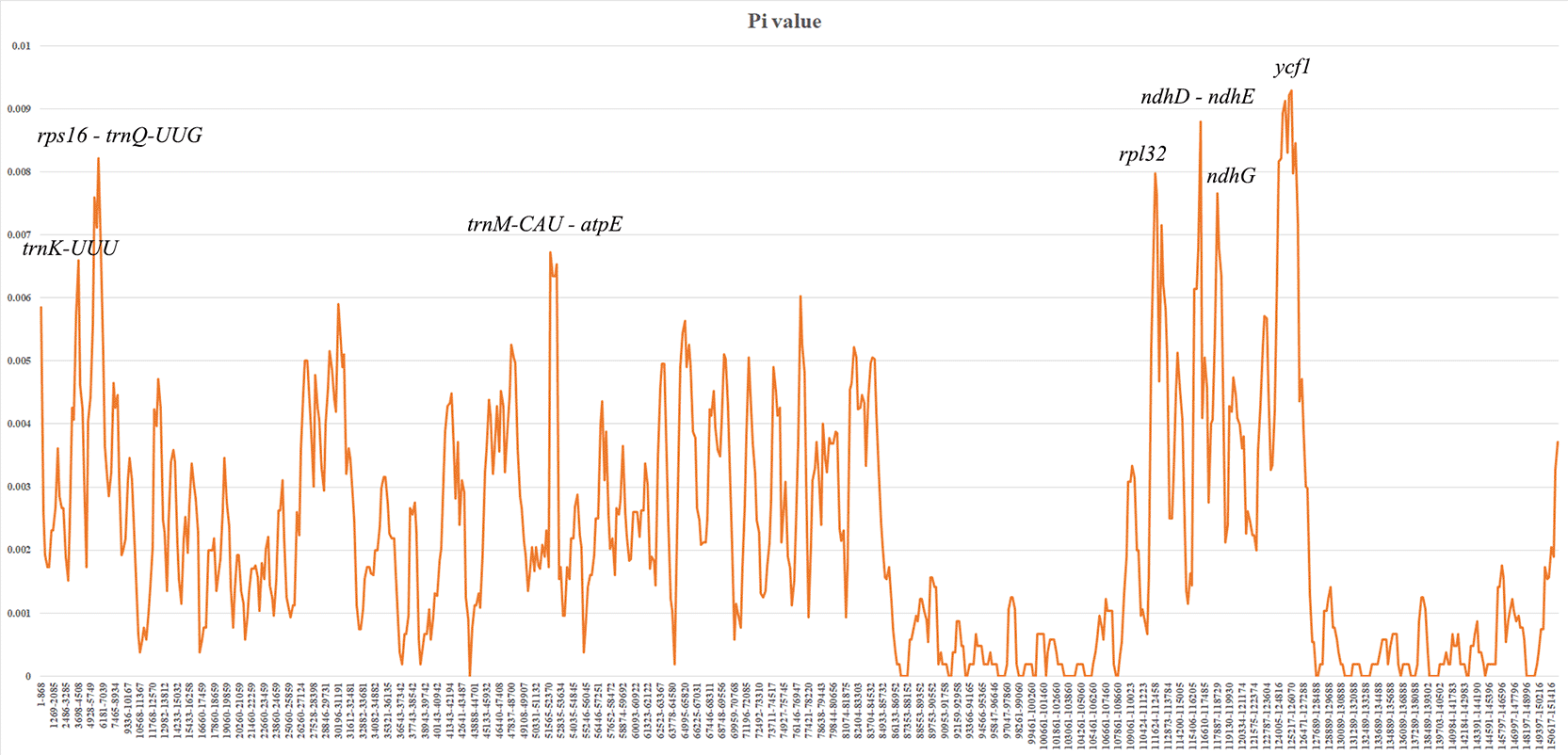
Nucleotide variability (Pi) is plotted along the genome sequence. The window length was set to 800 bp with a step size of 200 bp.
Overall, the present results show that D. helenae combines strong plastome structural conservatism with enough sequence divergence in a small number of highly informative loci to support phylogenetic placement and future species-level identification. Because D. helenae is an accepted Central Asian species of medicinal interest and several phytoecdysteroids have already been reported from it, the plastome resource generated here should be valuable for molecular authentication, population-level marker development, and future conservation-oriented studies of the genus in the region (Yusupova et al., 2022).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)