Keywords
Lepidium, chloroplast genome, phylogenetics, genetic diversity, ycf1 gene, plastome analysis, Brassicaceae
This article is included in the Genomics and Genetics gateway.
Lepidium, chloroplast genome, phylogenetics, genetic diversity, ycf1 gene, plastome analysis, Brassicaceae
The genus Lepidium L. (Brassicaceae), commonly known as pepperwort or peppercress, is a cosmopolitan taxon comprising between 175 and more than 260 species, depending on the circumscription and recent taxonomic revisions (Al-Shehbaz, 2012; De Lange et al., 2013; Koch and Mummenhoff, 2006; Bona, 2020). It is widely distributed across temperate and subtropical regions globally, with centers of diversity in Central Asia, Australia, New Zealand, and the Americas (German, 2014; Ilyinska, 2014; Rūrāne and Roze, 2019 ). Species within Lepidium inhabit diverse ecological niches, ranging from coastal zones to alpine environments, and display significant morphological diversity and adaptive specialization.
Historically, Lepidium has posed considerable challenges to systematists due to extensive morphological variation, frequent polyploidy, hybridization events, and the occurrence of both sexual and autogamous reproduction (Al-Shehbaz, 2012; De Lange et al., 2013; Dierschke et al., 2009). The genus is characterized by small, usually inconspicuous flowers often lacking petals, and fruits that are angustiseptate silicles with a single mucilaginous seed per locule, a feature aiding in long-distance dispersal (Koch and Mummenhoff, 2006). This combination of reproductive plasticity and dispersal capability has contributed to the broad distribution and complex taxonomy of the genus, with many species historically being grouped under broad or narrow concepts depending on regional traditions (German, 2014).
Recent molecular phylogenetic studies have provided significant insights into the intrageneric relationships within Lepidium, using both nuclear markers, such as ITS and ETS, and chloroplast markers, such as trnL-F , rbcL, and matK. Mummenhoff et al. (2001) established the first comprehensive chloroplast DNA phylogeny for the genus, revealing a major division into three lineages largely reflecting geographic distributions: the Eurasian lineage, including sections Lepia and Cardaria; the Australasian lineage, corresponding to Monoplocoidea; and a widespread clade comprising the remaining taxa. These findings were supported by more recent analyses using super-barcodes, including full plastome sequences, which demonstrated that complete chloroplast genomes can provide higher phylogenetic resolution in complex genera such as Lepidium (Zhou et al., 2023; Song et al., 2023).
Chloroplast genome, or plastome, sequencing has become a powerful tool for resolving phylogenetic relationships and evolutionary patterns within plant genera (Munavvarov et al., 2022). Complete plastome sequences offer robust phylogenetic signals due to their uniparental inheritance, low recombination rates, and conserved structure; however, they also contain evolutionary hotspots that may vary among lineages (Ergashov et al., 2026a; Ergashov et al., 2025a; Ergashov et al., 2025b; Ergashov et al., 2026b; Ergashov et al. 2026c; Jansen and Ruhlman, 2012). Plastome studies have been particularly useful in groups with intricate evolutionary histories, including cases of allopolyploidy and cryptic speciation, as observed in Lepidium (Dierschke et al., 2009; Mummenhoff et al., 2004).
Despite the increasing availability of Lepidium plastome data in public repositories such as NCBI, comparative analyses across a broad sampling of the genus remain limited. To address this gap, the present study undertakes a comprehensive plastome-wide comparative genomic analysis across 17 Lepidium species. This includes one newly sequenced species, Lepidium olgae, endemic to arid regions of Central Asia, particularly the Nuratau region of Uzbekistan, together with 16 publicly available plastome sequences. Lepidium olgae, a rarely studied species with ecological adaptations to xeric environments, provides a valuable addition to the genomic sampling of the genus, particularly for understanding plastome variation in Central Asian taxa (German, 2014; Song et al., 2023). The main objectives of this study are: (1) to characterize and compare the plastome structures, gene content, and sequence divergence among 17 Lepidium species; (2) to identify structural variations such as inversions, expansions and contractions of inverted repeats, and divergent hotspots; and (3) to infer phylogenetic relationships within the genus based on complete plastome data and evaluate their evolutionary implications for Lepidium taxonomy.
Through this comparative plastomic framework, we aim to contribute to a better understanding of chloroplast genome evolution in Lepidium, clarify phylogenetic relationships, and provide foundational data for future evolutionary and taxonomic studies in Brassicaceae.
Fresh leaves of Lepidium olgae Regel were collected from a wild population in the Nuratau Range, Khayotsoy, Uzbekistan, in May 2024 (40.508056° N, 66.723889° E; Figure 1). Species identification was carried out by N. Beshko at the National Herbarium of Uzbekistan, and representative voucher specimens were deposited in the National Herbarium of Uzbekistan (TASH) under voucher accession number TASH0007.

A, plant at the beginning of the growing season, top view, approximately 1300 m a.s.l., 3 April 2022; B, flowering individual, approximately 1300 m a.s.l., 6 May 2012; C–D, flowering plants, 14 April 2013; photographs by N. Beshko.
The collected leaf material was immediately dried in silica gel and stored at room temperature until DNA extraction. Total genomic DNA was extracted from dried leaf tissue using a Tiangen plant genomic DNA extraction kit following the manufacturer’s protocol. Sequencing libraries were prepared using a NEB library preparation kit for Illumina sequencing. The genomic DNA was fragmented to approximately 350 bp, followed by end repair, adapter ligation, and PCR amplification. Library quality and fragment size distribution were assessed using an Agilent 5400 system, and library concentration was subsequently determined. Qualified libraries were sequenced on an Illumina platform at Novogene Bioinformatics Technology Co.
Clean reads were assembled using NOVOPlasty (Dierckxsens et al., 2017). Gene annotation was performed in Geneious Prime v2025.1.2 using the chloroplast genome of Lepidium apetalum Willd. (GenBank accession NC_051540) as the reference. The annotation was manually checked and corrected, including verification of start and stop codons and exon–intron boundaries of protein-coding genes (Kearse et al., 2012). The circular chloroplast genome map was generated using OGDRAW (Greiner et al., 2019; Figure 2).
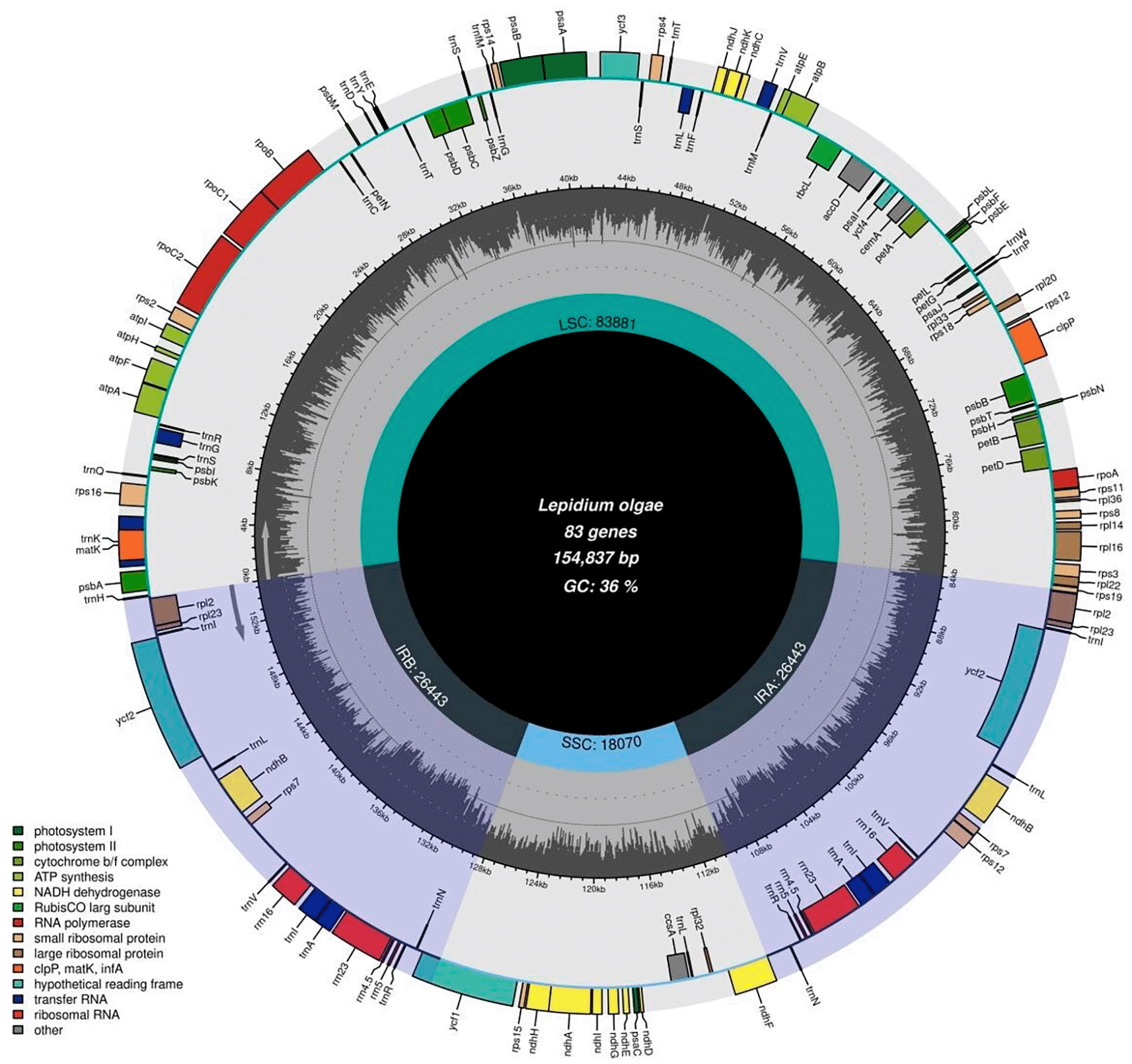
Genes located on the outer and inner circles are transcribed in opposite directions, respectively. The inner histogram represents the GC content across the genome. IRa and IRb indicate the inverted repeat regions, while LSC and SSC denote the large single-copy and small single-copy regions, respectively.
For comparative plastome analysis, 34 chloroplast genome accessions representing 17 Lepidium species were retrieved from NCBI GenBank ( Table 1). Complete chloroplast genome sequences were aligned using MAFFT v7.471/v7.520 (Katoh & Standley, 2013). Gene functions were classified into photosynthesis-related, self-replication-related, biosynthesis-related, and unknown-function categories.
| No. | Species name | NCBI accession number |
|---|---|---|
| 1 | Hornungia petraea | NC_049650 |
| 2 | Yinshania furcatopilosa | MK637818 |
| 3 | Yinshania henryi | MK637819 |
| 4 | Yinshania zayuensis | NC_062044 |
| 5 | Lepidium echinatum | NC_049660 |
| 6 | Lepidium perfoliatum | OQ644480 |
| 7 | Lepidium perfoliatum | MT880913 |
| 8 | Lepidium perfoliatum | ON598357 |
| 9 | Lepidium draba | NC_077506 |
| 10 | Lepidium appelianum | NC_077510 |
| 11 | Lepidium chalepense | NC_077508 |
| 12 | Lepidium chalepense | ON598368 |
| 13 | Lepidium olgae | PV605702 |
| 14 | Lepidium latifolium | ON598362 |
| 15 | Lepidium latifolium | ON598361 |
| 16 | Lepidium latifolium | ON598359 |
| 17 | Lepidium sp. XJ-151 | OQ644481 |
| 18 | Lepidium cartilagineum | NC_077509 |
| 19 | Lepidium didymum | OY986990 |
| 20 | Lepidium meyenii | NC_034363 |
| 21 | Lepidium meyenii | MT430983 |
| 22 | Lepidium meyenii | KY231152 |
| 23 | Lepidium cordatum | NC_077507 |
| 24 | Lepidium apetalum | NC_051540 |
| 25 | Lepidium apetalum | OR941701 |
| 26 | Lepidium apetalum | PP234589 |
| 27 | Lepidium ruderale | NC_077504 |
| 28 | Lepidium ruderale | ON598356 |
| 29 | Lepidium sativum | NC_047178 |
| 30 | Lepidium sativum | MK637743 |
| 31 | Lepidium virginicum | NC_009273 |
| 32 | Lepidium ferganense | ON598364 |
| 33 | Lepidium ferganense | OQ644478 |
| 34 | Lepidium ferganense | NC_077505 |
The structural characteristics of the chloroplast genomes, including the large single-copy region, small single-copy region, and inverted repeat regions, were compared among the sampled Lepidium species. Expansion and contraction of the inverted repeat regions were analyzed using IRscope (Amiryousefi et al. 2018) and manually checked in Geneious Prime v2025.1.2. The junction regions were defined as JLB, the junction between LSC and IRb; JSB, the junction between SSC and IRb; JSA, the junction between SSC and IRa; and JLA, the junction between LSC and IRa. Variation in these boundary regions was recorded to evaluate plastome structural diversity within Lepidium.
Nucleotide diversity (Pi) was calculated using DnaSP v6.12.03 (Rozas et al., 2017). A sliding-window analysis was performed with a window length of 800 bp and a step size of 200 bp to identify highly variable regions across the chloroplast genomes. Regions showing elevated Pi values were considered potential molecular markers for phylogenetic and DNA barcoding studies.
For phylogenetic analysis, 34 complete chloroplast genome sequences were used, including 30 accessions representing 16 named Lepidium species and one unidentified Lepidium accession, together with four outgroup accessions from Hornungia and Yinshania ( Table 1). Maximum likelihood analysis was performed in RAxML with 1,000 bootstrap replicates (Stamatakis, 2014). The GTR + G model was selected using jModelTest v2.1.4 under the Akaike Information Criterion (Darriba et al., 2012).
The chloroplast genomes of the 17 analyzed Lepidium species showed the typical circular quadripartite structure of angiosperm plastomes, consisting of a large single-copy (LSC) region, a small single-copy (SSC) region, and a pair of inverted repeat regions, IRa and IRb ( Figure 2). The total plastome size ranged from 153,132 bp to 154,982 bp, indicating a high level of structural conservation across the genus. The chloroplast genome of Lepidium olgae was 154,837 bp in length. All analyzed plastomes encoded 127 unique genes, including 82 protein-coding genes, 37 transfer RNA genes, and 8 ribosomal RNA genes. These genes were mainly associated with photosynthesis, transcription, translation, and essential metabolic processes. Gene content, gene order, and overall genomic organization were highly conserved among the examined species, and no major rearrangements were detected. This pattern agrees with previous plastome studies in Brassicaceae and other angiosperms, where conserved quadripartite structure, limited gene loss, and stable genome organization have commonly been reported (Jansen et al., 2007; Wicke et al., 2011; Daniell et al., 2016; Huang et al., 2022; Dekhkonov et al., 2025; Tojiboeva et al., 2025; Nikitina et al., 2025).
Comparative analysis of the LSC, SSC, and IR junctions revealed only minor variation among the 17 Lepidium chloroplast genomes ( Figure 3). At the JLB boundary, located between LSC and IRb, the rps19 gene was partially duplicated into the IRb region in most species, with slight differences in the length of the overlapping segment. At the JSB boundary, the ndhF gene consistently extended slightly into the IRb region. The JSA boundary showed limited variation, mainly related to the position and length of the ycf1 fragment extending into IRa, while the JLA boundary was highly conserved, with adjacent genes such as rpl2 and psbA maintaining stable positions. These minor expansions and contractions of IR regions are likely lineage-specific microstructural changes rather than major plastome reorganizations. Similar IR boundary shifts have been widely reported across angiosperms and are considered one of the main contributors to plastome size variation (Zhu et al., 2016; Wang et al., 2008). The low variability of IR regions also supports the idea that duplicated regions are under stronger evolutionary constraint, probably due to copy-correction mechanisms and reduced substitution rates (Birky and Walsh, 1992; Wicke et al., 2011; Smith, 2015).

JLB represents the junction of LSC and IRb, JSB indicates the junction of SSC and IRb, JSA denotes the junction of SSC and IRa, and JLA signifies the junction of LSC and IRa.
Sliding-window analysis of nucleotide diversity showed heterogeneous sequence variation across the Lepidium plastomes ( Figure 4). Pi values ranged from approximately 0.002 to 0.047, indicating generally low to moderate sequence divergence among species. Several highly variable regions with Pi values above 0.03 were detected, mostly in intergenic spacers and several coding regions, including trnQ–psbK, trnD, trnL–trnF, psbJ–psbL, rpl14–rpl16, and rpl32–ccsA. The highest nucleotide diversity was observed in the ycf1–trnN region, where Pi values reached approximately 0.045–0.047. In contrast, IR regions showed distinctly lower nucleotide diversity than the LSC and SSC regions, reflecting the conserved nature of duplicated plastome regions.
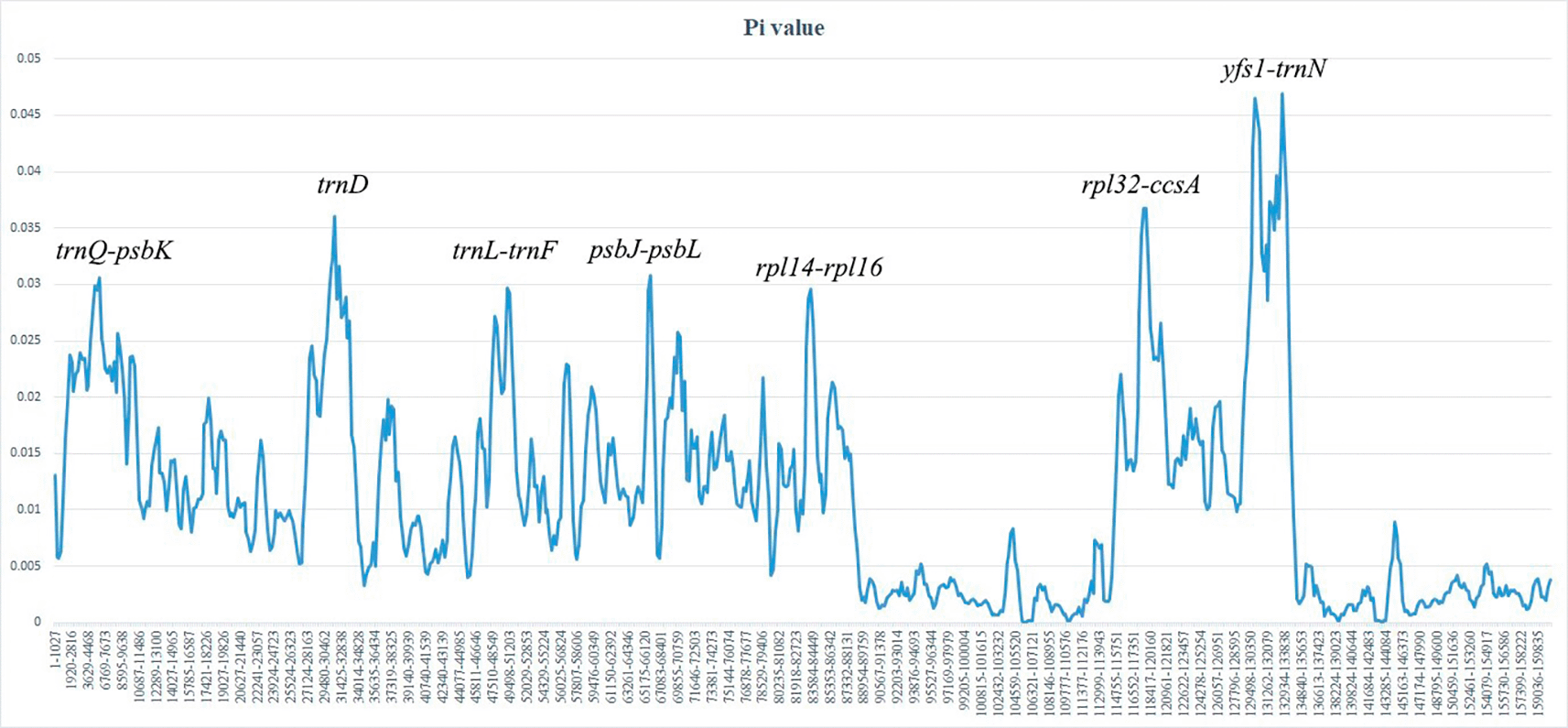
Nucleotide variability (Pi) is plotted along the genome sequence. The window length was set to 800 bp with a step size of 200 bp.
The elevated variability of the ycf1–trnN region is consistent with previous studies showing that ycf1 is one of the fastest-evolving regions in angiosperm plastomes and one of the most promising plastid DNA barcodes for land plants (Jansen et al., 2007; Dong et al., 2015; Kuang et al., 2011). In addition, non-coding regions such as rpl32–trnL-UAG , trnL–trnF, and psbJ–psbL showed considerable variation, which is expected because intergenic spacers are generally under weaker functional constraint and accumulate substitutions more rapidly than coding regions (Daniell et al., 2016; Shaw et al., 2007). Comparable patterns of divergence have also been reported in plastome-wide analyses of Cardamine, Brassica, and other Brassicaceae genera (Huang et al., 2022; Ergashov et al., 2026a). Therefore, the hypervariable loci identified in the present study may serve as useful molecular markers for species delimitation, phylogenetic reconstruction, and fine-scale phylogeographic studies in Lepidium.
Phylogenetic analysis based on complete chloroplast genome sequences strongly supported the monophyly of Lepidium ( Figure 5). The maximum likelihood tree resolved major clades with bootstrap support values ranging from 67% to 100%, indicating that complete plastome data provide a robust phylogenetic signal for this genus. Multiple accessions of L. chalepense formed a strongly supported clade, while accessions of L. latifolium, L. meyenii, L. apetalum, L. ruderale, L. sativum, and L. ferganense clustered according to species identity. This pattern suggests high genetic consistency among accessions and confirms the usefulness of whole chloroplast genomes for resolving species-level relationships in Lepidium. The inclusion of related genera further supported the clear separation of Lepidium from closely related Brassicaceae lineages.

Phylogenetic relationships were inferred from complete chloroplast genome sequences. Bootstrap support values are shown at the nodes. Related genera of Brassicaceae, including Yinshania and Hornungia, were used as outgroups.
.
The topology of the plastome tree broadly reflected geographic structuring within the genus. The clustering of L. perfoliatum, L. draba, L. appelianum, and L. chalepense supports the presence of a shared Eurasian lineage, in agreement with earlier chloroplast DNA phylogenies (Mummenhoff et al., 2004; Mummenhoff et al., 2001). Similarly, South American taxa such as L. meyenii and L. didymum formed a distinct clade, suggesting independent diversification after long-distance dispersal events. The distinct placement of L. echinatum from Australia and the Central Asian endemic L. olgae highlights the importance of geographic isolation in lineage divergence. These results are consistent with biogeographic hypotheses suggesting multiple intercontinental dispersal events in Lepidium during the Neogene (German, 2014; Mummenhoff et al., 2001).
The phylogenomic pattern observed here suggests that diversification in Lepidium has occurred mainly through geographic radiation and ecological differentiation rather than large-scale plastome restructuring. Mucilaginous seed coats, which are characteristic of Lepidium, may have promoted long-distance dispersal through epizoochory and water-mediated transport, facilitating colonization of new habitats (Rūrāne and Roze, 2019; Mummenhoff et al., 2001 ). Together with predominantly autogamous reproductive systems, this dispersal strategy may have supported rapid establishment in newly colonized areas while maintaining species-level genetic cohesion. Autogamy is also associated with reduced recombination and lower within-population genetic diversity, which may partly explain the moderate plastome divergence observed in the genus (Glémin et al., 2006; Wright et al., 2008).
Overall, the present results demonstrate that chloroplast genomes of Lepidium are structurally conservative, with only minor IR boundary shifts and moderate sequence divergence. At the same time, several highly variable regions, particularly ycf1–trnN, provide useful candidates for molecular marker development. The well-supported phylogenetic topology confirms the value of whole plastome data for clarifying taxonomic boundaries and evolutionary relationships within Lepidium. However, because the chloroplast genome represents a single maternally inherited locus, future studies incorporating nuclear genomic data will be necessary to resolve possible hybridization, reticulate evolution, and polyploidization events in the genus (Birky, 2001; Mummenhoff et al., 2004). Divergence-time estimation and comparative Ka/Ks analyses of rapidly evolving genes such as ycf1 may further clarify whether the detected hotspots reflect relaxed purifying selection or lineage-specific adaptive evolution (Yang and Nielsen, 2000).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)