Keywords
Camelidae, climatic variables, density effects, guanaco, population regulation, population growth rate
Camelidae, climatic variables, density effects, guanaco, population regulation, population growth rate
In order to understand population dynamics and optimize the management of wildlife populations it is important to identify how groups are affected by environmental factors and their own density, particularly in species living in extreme environments. Mechanisms that regulate populations of the guanacos (Lama guanicoe) are poorly known; this species is heavily exploited in the Patagonian steppe, and its numbers and distribution have diminished significantly during the last century because of grazing conflicts with a sheep-based society, and overhunting (the guanaco’s historical distribution has been reduced by 75% in Chile and Peru, and by 60% in Argentina)1. The guanaco is in the Least Concern IUCN Red List category (http://www.iucnredlist.org/, downloaded on 11 September 2013) but it has been included in Appendix II of CITES 2013 (http://www.cites.org/eng/app/2013/E-Appendices-2013-06-12.pdf).
No population of any species can grow indefinitely, and population checks based upon different processes restrict population size and/or geographic distribution; these processes are either density-stabilizing or density-limiting2, the latter being independent of population size. Stabilization results from density-dependence, with a regulatory effect that varies in intensity with the size or density of the population itself. However, density-dependent processes are also affected by environmental conditions, and many wildlife population dynamic and management models include the effects of climatic covariates (e.g., Dennis & Otten, 2000; Colchero et al., 2009)3,4. In the case of the Ricker model (one of the most simple and most used population models), Corani and Gatto (2007)5 proposed an original way of incorporating climatic covariates: they were included as affecting the population growth rate.
The guanaco is one of the two extant wild South American camelids, and ranges from Northern Peru through Chile, and across Patagonia to southern Argentina and Chile, reaching Tierra del Fuego. It is found from sea level to nearly 4500 m on the Andes mountain range, and occupies a wide variety of habitats from hardpan deserts to scrublands to grasslands6.
Although there are several studies on the population regulation processes in mammals in general7,8, and in ungulates in particular9–12, there are very few studies in wild South American camelids that analyze population growth regulation. There have been some studies on populations of vicuña (Vicugna vicugna)13–15 and some on populations of guanacos16–18; however none of these studies considered the direct effect of density and/or climate on population growth rate.
Thus, we tested the hypothesis that population density and environmental covariates, such as average winter temperature and average annual precipitation, affect the growth rate of the guanaco population from the island of Tierra del Fuego, Chile.
We analyzed the guanaco population of the “Cameron” ranch (-53.9 S, -69.3 W), with an area of the 2000 km2 located in the Southern region of the Tierra del Fuego island, Chile. The altitude range is 0–300 m above sea level, and the ranch is a mosaic of steppe and forest biomes; the latter is a deciduous forest and the steppe is composed of meadows, peats and prairies. The guanaco is the dominant herbivore, with the exception of sheep (the dominant domestic species) with densities that have fluctuated in the last decades between 11 and 23 sheep/km226. Based on metabolism studies19, the guanaco/sheep equivalence factor is 1.65 (i.e., each sheep is equivalent to 0.61 guanacos), close to the values of 1.5 and 1.8 sheep per guanaco proposed by Raedeke (1978)20 based on diet habits. In non-forested areas (the Patagonian steppe) the climate is characterized by an average annual precipitation of 200–400 mm, while in the forested areas the average precipitation fluctuates between 400–600 mm per year20. The average annual temperature is 6.5°C and the average winter temperature is 2.2°C.
Guanacos were counted for 34 consecutive years between 1977 and 2012 (except 1986 and 1996), using the transect method with a variable width, and a maximum of 1000 m to each side of the transect from 1977 to 2000, and with a fixed width band from 2001 to 2012 (the latter with a maximum of 500 m to each side of the transect)21–25. The sampling period was carried out in the autumn and lasted approximately 7 days between 10:30 and 19:00 h, with two observers in two 4x4 vehicles going over the main, secondary and local roads at a maximum speed of 40 km/h. Each road was covered only once, and in addition to individual guanaco counts, the following were recorded: weather conditions, time, distance (km) from the starting point, GPS coordinates, observation distance from the transect (m), an estimate of the angle to the animal’s position, and – when the animals were observed in groups – the number of individuals, the type of social group, and its structure (sex, and the age class: newborn, juvenile and adult). Observations were done with model Trophy of Bushnell® 10x42 binoculars, and the GPS was a Garmin model eTrex Vista. Observation distances were made by naked eye, but the four observers had been trained in this technique using laser telemeters, GPS and the vehicles’ odometers to calibrate the estimated distances. The road network and all geo-referenced observations were processed with the Arc View 9.3 Geographical Information System (SIG), and transferred using program Map Source®. The cartography was kindly provided by the Chilean “Servicio Agrícola y Ganadero” (SAG).
The population size was estimated as given in Soto26 which was based in the method described by Raedeke (1978)20, given by:
where N is the total population to be estimated, A is the total study area, n is the total number of animals counted, x is the total transect distance covered (m) rounded to one meter, and y is the average of the perpendicular distance (m) from the transect to the animals counted (the factor of 2 is included to consider that there is one band to each side of the transect). The variance (S2) was estimated by (2), with p = n/N, and used to estimate the 95% upper and lower confidence intervals.
In addition to the direct density-dependence, we evaluated the possible impact of two environmental covariates on guanaco population dynamics: average annual precipitation and winter temperature (as the average of months of June, July and August). We used the 25 years (1977–2002) precipitation and temperature time series of the CRU TS 2.1 database, compiled by the Tyndall Centre, Climatic Research Unit, School of Environmental Sciences of the University of East Anglia, United Kingdom (http://www.cru.uea.ac.uk/cru/data/hrg.htm). As the CRU TS 2.1 data ended in 2002, we completed that time series for 2003 to 2012 from the closest meteorological station to the Cameron ranch: Punta Arenas (Chile); this data was downloaded from the Internet site of the Meteorological Service of Chile (http://www.meteochile.gob.cl/).
We estimated the population growth rate (as measured by the finite net rate of increase, λ) for each of the 36 years of the guanaco data (though population values were not available for years 1986 and 1996, and were linearly interpolated) by fitting a three age-class (newborns, juveniles and adults) matrix model to the field population estimates; as the transition matrix is a female only matrix we summed the three age classes and multiplied by two (in this population there is a 50% sex-ratio) to carry out the fit with the total field population. The fit was carried out in an Excel® spreadsheet using the Solver tool, and the sum of squares (SSQ) goodness of fit criterion was used (details can be found in Rabinovich and Zubillaga, 201227); this process resulted in a set of 36 population stage-structured matrices. From each matrix we calculated the largest positive eigenvalue28 as an estimate of λ; with the PopTools add-in, an Excel program developed by Greg Hood (http://www.cse.csiro.au/poptools/).
We checked the relationship between the population growth rate (as measured by the finite net rate of increase, λ) and population size (in units of natural logs) and/or climate using a stepwise regression analysis with λ as dependent variable, and total guanaco population (in natural logs), annual mean precipitation (mm/year), and winter mean temperature (°C) as independent variables. For the climatic covariates we also evaluated the effect of time lags (T), with T = 1 to 7 years for precipitation, and T = 1 year for winter temperature; the lags were applied by averaging the previous T lagged years, as suggested by Shaw et al. (2012)14. These regressions were carried out using the statistical package Statistica (StatSoft, 2009; http://www.statsoft.com/). Since it is necessary to corroborate that there is no collinearity (significant correlations among the predictor variables) for multiple regressions analysis, we carried out correlation analysis between independent variables before the stepwise analysis. A p value < 0.05 was considered significant.
The table in Supplementary Table 1 shows the guanaco abundance data for the 36 years of sampling, the estimated values of the population rate of growth (λ) from the 36 population transition matrices, and the values of climatic variables used in the regression analysis.
The precipitation covariates with 1 to 7 year lags showed a statistically significant Pearson's correlation coefficient (p < 0.05), while the other independent variables didn’t show a statistically significant correlation. Thus, for the stepwise analyses we chose one of these correlated covariates (precipitation with lag T = 4). The results indicated that the total population (LnNtot) was the only statistically significant variable (Table 1), and the model was statistically significant (p < 0.030 and F(5, 30) = 2.885). Table 1 shows both the standardized and the raw regression coefficients, with their corresponding standard errors. The magnitude of these standardized regression coefficients allows comparison of the relative contribution of each independent variable in the prediction of the dependent variable.
The results show the standardized regression coefficients (Sz. Reg. coeff.), their standard errors (Std.E. SRC), the raw regression coefficients (Reg. Coeff.), their standard errors (Std.E. RC) and t-test values (t-stud.).
We carried out another regression analysis using λ as the dependent variable and only population size (in natural logs, LnNtot) as an independent variable, in order to compare it with the full model (population size and climatic covariates as independent variables). Table 2 shows the result of this second regression analysis, which was highly significant (p < 0.0018, r = 0.502); the regression equation was λ = 1.6373–0.0552 (±0.0163) LnNtot; a comparison of both regressions using a two-way ANOVA test indicated that there was no statistical significant differences between them (F = 0.803, p = 0.5329).
Interpretation of the statistical parameters as in Table 1. The results show the standardized regression coefficients (Sz. Reg. coeff.), their standard errors (Std.E. SRC), the raw regression coefficients (Reg. Coeff.), their standard errors (Std.E. RC), and t-test values (t-stud.).
| Sz.Reg. coeff. | Std.E. SRC | Reg. Coeff. | Std.E. of RC | t-stud.(30) | p-value | |
|---|---|---|---|---|---|---|
| Intercept | - | - | 1.6373 | 0.1624 | 10.0825 | 9.45E-12 |
| LnNtot | -0.5024 | 0.1483 | -0.0552 | 0.0163 | -3.3879 | 0.00179 |
The negative slope is an indication of a density-dependent process, since when the population increases the per capita population growth rate decreases (Figure 1). The intercept of the regression line at λ = 1 (population at equilibrium) results in a population size of 103,000 guanacos (51 guanaco/km2), which can be considered as an estimate of the carrying capacity of the Cameron ranch for this guanaco population.
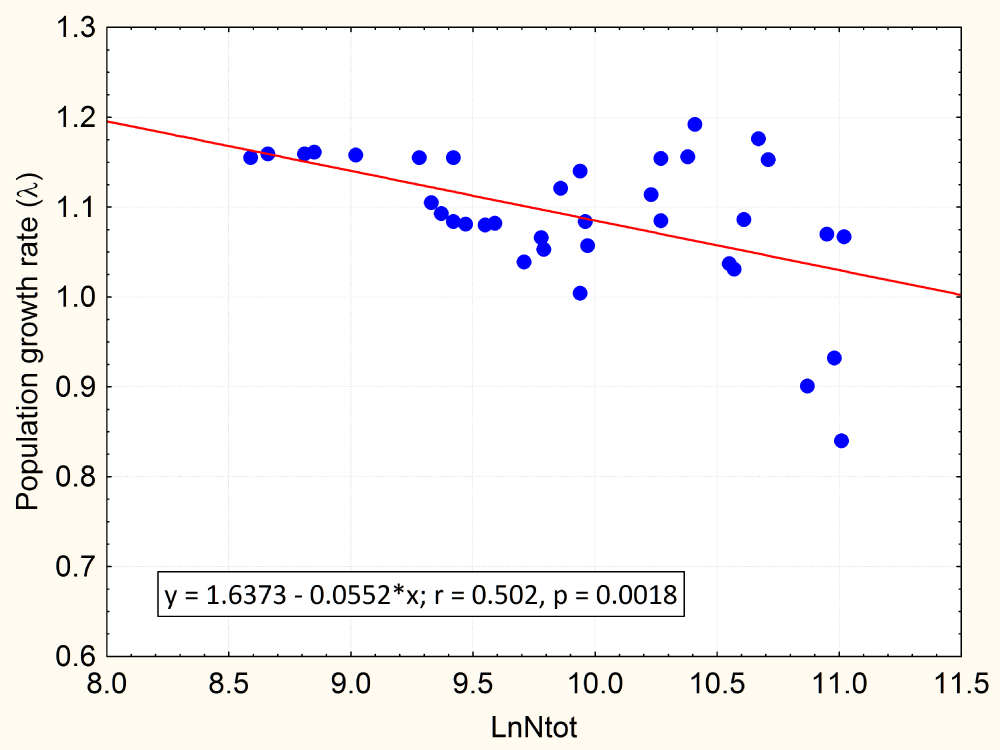
On the x-axis the guanaco population from the Cameron ranch (Tierra del Fuego, Chile) is given in a natural logarithmic scale. The values of λ represent the finite population growth rate.
We were not able to confirm the expected effect of climatic covariates on the population rate of growth in the guanaco population of the Cameron ranch. We expected such an effect because Sarno et al. (1999)18 found a relationship between winter weather and guanaco yearling’s survival, and Rey et al. (2012)29 recorded an effect of drought on guanaco fecundity. Thus we anticipated that as winter temperature and/or annual precipitation decrease, the population growth rate should also decrease. Our negative results are even more surprising because in the last 10–12 years the guanaco population fluctuated remarkably, suggesting some sort of interaction between population density and climatic covariates. It is well known that environmental effects are more important when the population size is near the carrying capacity, particularly in large mammals (as the guanaco) characterized by low reproductive rates, long life-spans, and populations that are resource-limited (features typical of species referred to as the “K-selected” species)30. The difference between our results and that of Sarno et al. (1999)18 may reflect that these authors used the average winter snowfall while we considered average winter temperature.
Taper & Gogan (2002)31 carried out a study with the Yellowstone elk similar to our analysis, although the weather variables were included as covariates within a dynamic model (exponential, Ricker and Gompertz models). They found that spring precipitation had a positive regression coefficient, a result opposite to our conclusion for the guanaco population.
Our conclusion is that in order to test the effects of climatic covariates on population regulation of large ungulates, the use of a population dynamic model is recommended, for they may be more sensitive to the interaction between density-dependent processes and weather variables, than a simple regression between them and population growth rate. For example, our negative results do not conform to what was obtained by Shaw et al. (2012)14 with respect to a very phylogenetically related camelid species: the vicuña. Using a logistic model fitted to 31 years of data these authors found that rainfall had a highly significant effect on population size with a time lag of 4 years, while in our guanaco regression analysis, no climatic covariate was statistically significant.
We conclude that, opposite to what we expected based on the bibliography of ungulate’s population dynamics, weather variables do not seem to influence the density-dependent population growth rate process.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)