Chakraborty S, Rao BJ, Asgeirsson B and Dandekar AM. Correlating the ability of VP24 protein from Ebola and Marburg viruses to bind human karyopherin to their immune suppression mechanism and pathogenicity using computational methods [version 2; peer review: 2 approved with reservations]. F1000Research 2014, 3:265 (https://doi.org/10.12688/f1000research.5666.2)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
Correlating the ability of VP24 protein from Ebola and Marburg viruses to bind human karyopherin to their immune suppression mechanism and pathogenicity using computational methods
[version 2; peer review: 2 approved with reservations]
Sandeep Chakraborty1,2, Basuthkar J. Rao2, Bjarni Asgeirsson3, Abhaya M. Dandekar1
Sandeep Chakraborty1,2, Basuthkar J. Rao2, Bjarni Asgeirsson3, Abhaya M. Dandekar1
1Plant Sciences Department, University of California, Davis, CA 95616, USA 2Department of Biological Sciences, Tata Institute of Fundamental Research, Homi Bhabha Road, Mumbai, 400 005, India 3Science Institute, Department of Biochemistry, University of Iceland, Dunhaga 3, IS-107 Reykjavik, Iceland
This article is included in the Ebola Virus collection.
Abstract
Immune response suppression is crucial for viral invasion. The protein VP24 is pivotal in achieving this in Ebola, although interestingly the mechanism of immune suppression is different in the closely related Marburg virus. Here, we illustrate that a possible molecular basis for this difference emanates from two alpha helical structures (α5 and α6) in VP24 involved in binding human karyopherin (KPNA) (PDBid:4U2X), wherein the Ebola and Marburg viruses have distinctly different charged properties in α5. α6 is absent in Marburg, and has a different hydrophobic moment in the Reston Ebola (REBOV) species, which is surprisingly non-pathogenic in humans. Based on the hypothesis that REBOV is not immunosuppressive, which is in turn is due to its inability to bind KPNA, we show by docking KPNA to the REBOV VP24 that the single amino acid substitution R140S is responsible for this difference between REBOV and Zaire Ebola strains. Such a scenario of getting a virulent REBOV through a single mutation is particularly worrisome, since the REBOV, once found only in monkeys, has been recently detected in pigs. We also reiterate the potential of using these helices as potential epitopes for generating protective antibodies against Ebola.
Corresponding author:
Sandeep Chakraborty
Competing interests:
No competing interests were disclosed.
Grant information:
AMD wishes to acknowledge grant support from the California Department of Food and Agriculture PD/GWSS Board. BJ acknowledges financial support from Tata Institute of Fundamental Research (Department of Atomic Energy). Additionally, BJR is thankful to the Department of Science and Technology for the JC Bose Award Grant. BA acknowledges financial support from the Science Institute of the University of Iceland.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
In this version, based on the suggestion of a reviewer (Dr McIntosh), we have docked the modelled structure of VP24 from Reston Ebola (erVP24) virus to the structure of mouse KPNA (PDBid:1Y2AC) (there are no known models of KPNA from other non-human primates that are susceptible to Reston Ebola virus). However, inspite of sequence difference (50.3\% identity and 77.8\% similar) between human and mouse KPNA, the residues that interact with erVP24 is the same. However, Reston Ebola is known to be non-virulent in mice, although viral replication does occur (de Wit E, et. al.). Virulence in mice is caused only by a STAT1 knockout, corroborating the inability of erVP24 to suppress immune response, as hypothesized by us as a major cause for its non-pathogenicity. The title of the article has been changed, Dataset 1 has been updated and a new figure (fig 5) has been added.
In this version, based on the suggestion of a reviewer (Dr McIntosh), we have docked the modelled structure of VP24 from Reston Ebola (erVP24) virus to the structure of mouse KPNA (PDBid:1Y2AC) (there are no known models of KPNA from other non-human primates that are susceptible to Reston Ebola virus). However, inspite of sequence difference (50.3\% identity and 77.8\% similar) between human and mouse KPNA, the residues that interact with erVP24 is the same. However, Reston Ebola is known to be non-virulent in mice, although viral replication does occur (de Wit E, et. al.). Virulence in mice is caused only by a STAT1 knockout, corroborating the inability of erVP24 to suppress immune response, as hypothesized by us as a major cause for its non-pathogenicity. The title of the article has been changed, Dataset 1 has been updated and a new figure (fig 5) has been added.
Viruses from the family Filoviridae are negative-stranded RNA viruses having a filamentous shape1. The first member of this family (Marburg) was discovered in 19672, while the Ebola virus was first discovered in 19763. Public attention has been drawn to this rare, but deadly disease4 ever since the current outbreak in West African countries threatened to rapidly deteriorate into a full-blown epidemic5,6. Both viruses cause haemorrhagic fever by quickly suppressing innate antiviral immune responses7. However, quite surprisingly, the Reston Ebola (REBOV) strain, first identified in monkeys that were imported into Reston in the United States from the Philippines8, is non-pathogenic in humans9,10.
Previously, we have characterized α-helical (AH) structures in Ebola proteins using PAGAL11, and demonstrated that the AHs with characteristically unique feature values are involved in critical interactions with host proteins12. We showed that the AH from Ebola virus membrane fusion subunit GP213, which is disrupted by a neutralizing antibody derived from a human survivor of the 1995 Kikwit outbreak14, has a very large hydrophobic moment compared to other AHs in Ebola proteins12. Similarly, another AH with the highest proportion of negatively charged residues is the binding site of the human karyopherin (KPNA) to the Zaire Ebola (ZEBOV) virus VP24 (ezVP24) protein15.
In spite of sharing a common ancestry16, Marburg and Ebola have different antigenicity of the virion glycoprotein17. Furthermore, the mechanism of immunosuppression is different in these viruses18. These differences are probably the reason for the reduced mortality observed in Marburg outbreaks. In Ebola, the crucial role of host immune system evasion is accomplished by two proteins: VP35 and VP2419. Ebola VP24 inhibits interferon (IFN) signaling by hindering the nuclear accumulation of tyrosine-phosphorylated STAT1 by binding KPNA20,21. In contrast, the Marburg virus abrogates the host immune response by inhibiting IFN-induced tyrosine phosphorylation of STAT1 and STAT218 via a moonlighting function matrix protein, VP4022. Specifically, ezVP24 binds KPNA via two AHs (α5 and α6)15. In Marburg VP24 (mVP24), α5 has distinctively different properties (not easily identified by a sequence or structural alignment), while α6 is just a small turn23. This explains why mVP24 is not immunosuppressive.
We investigated these AHs in VP24 from the REBOV strain (erVP24). While α5 in erVP24 was similar to that in ezVP24, α6 in erVP24 had different properties caused by the presence of a serine in the place of arginine (S140R). We modeled the apo erVP24 (PDBid:4D9OA) using the ezVP24 in complex with KPNA as a template (PDBid:4U2X) by SWISS-MODEL24, and then docked KPNA to this structure using DOCLASP25. The docked structure helped visualize the ability of Arg140 in ezVP24 to make the correct electrostatic interaction with two glutamic acids, one residing on α5 in VP24, and the other in KPNA. The effect of single mutations in modulating virulence has been well established26–28. However, our methodology provides a more rational way of finding such critical residues. The possibility of a REBOV mutant gaining immunosuppressive capabilities is particularly disconcerting since the isolation of the REBOV strains from pigs29–31. We also highlight the possibility of using α5 and α6 from VP24 as epitopes for generating antibodies32 or designing compounds and peptides to inhibit protein-protein interaction33.
Materials and methods
AHs in proteins were identified using DSSP34. These AHs were then analyzed using PAGAL11. Briefly, the Edmundson wheel is computed by considering a wheel with centre (0,0), radius 5, first residue coordinate (0,5) and advancing each subsequent residue by 100 degrees on the circle, as 3.6 turns of the AH makes one full circle. We compute the hydrophobic moment by connecting the center to the coordinate of the residue and giving it a magnitude obtained from the hydrophobic scale obtained from35. These vectors were then added to calculate the final hydrophobic moment. The color coding for the Edmundson wheel was as follows: all hydrophobic residues were colored red, while hydrophilic residues were colored in blue: dark blue for positively charged residues, medium blue for negatively charged residues and light blue for amides.
The protein structures used in the current work were all identified using the PDBid, and are available at www.rcsb.org. We used the SWISS-MODEL program to model the erVP24 (PDBid:4D9OA) structure using the ezVP24 (PDBid:4U2XA) in complex with KPNA as template. See 4D9OA4U2XA.pdb in Dataset 1 Note the residue numbering is not conserved by SWISS-MODEL. For example, Glu113 in PDBid:4D9OA corresponds to Glu97 in PDBid:4D9OA4U2XA. We used DOCLASP25 to dock KPNA to the modelled structure of erVP24 (See Pymol script ‘dockingKPNAtoRestonVP24.p1m’ for human KPNA and ‘RESTONVP24mouse.p1m’ for mouse KNPA in Dataset 1). ‘4U2XA.4U2XD.maxdist.out.sort’ in Dataset 1 lists the closest atoms of the residues of VP24 (PDBid:4U2XA) that make contact with human karyopherin (PDBid:4U2XD), sorted based on distances.
All protein structures were rendered by PyMol (http://www.pymol.org/). The sequence alignment was done using ClustalW36. The alignment images were generated using SeaView37. Protein structures were superimposed using MUSTANG38.
Results and discussion
Dataset 1.Version 2. Data used for SCALPEL search methodology to identify plant alpha helical - antimicrobial peptides in the PDB database.
list.plants.txt: list of PDB IDs resulting from querying the PDB database with the keyword ‘plant’. ALPHAHELICES.zip: DSSP analysis of proteins listed in list.plants.txt to identify alpha helices. RawDataHelix.txt: PAGAL analysis of alpha helices listed in ALPHAHELICES.zip. HTH: Set of all pairs of alpha helices connected with a short (<five residues) loop. RESTONVP24mouse.p1m’ is the pymol script for viewing the docked mouse KNPA to the modelled erVP24. blastkpna.png shows the different organisms whose KPNA structures have been solved.
Differences in α5 in Ebola and Marburg viruses: explaining why Marburg VP24 is not immunosuppressive
ezVP24 has a 39.6% identity (73.8% similar) with mVP24 (Figure 1a), and there is significant structural homology among VP24 proteins from different strains of Ebola and Marburg (Figure 1b). Yet, the mechanism of immune response suppression is different in these viruses from the Filoviridae family18. ‘Reasons why Marburg virus VP24 is not immunosuppressive remain elusive’23. Therefore, we sought to investigate the differences in residues involved in binding KPNA in the ezVP24 and mVP24.
Figure 1. Sequence and structural homology between VP24 proteins from different strains of Ebola and Marburg.
(a) EbZaire: Zaire Ebola, EBSudan: Sudan Ebola, EBReston: Reston Ebola, Mar-Musoke: Marburg Musoke. Multiple sequence alignment was done using ClustalW. Note, that the numbering used by ClustalW is not consistent with the real numbering of the VP24 residues. (b) Structural alignment of PDBid:4M0QA (Ebola Zaire Apo, in red), PDBid:4U2XA (Ebola Zaire complexed, in green), PDBid:4D9OA (Ebola Reston Apo, in blue), PDBid:3VNEA (Ebola Sudan Apo, in yellow) and PDBid:4OR8A (Marburg Musoke Apo, in orange). Structural alignment was done using MUSTANG38. (c) Helices involved in binding human karyopherin (α5 and α6 in magenta). Note, that the α5 is not a helix in Marburg VP24 (PDBid,4OR8A, in orange), but just a small turn.
ezVP24 binds KPNA via two AHs (α5 and α6), residues on loops and a Lys on a β-sheet (Table 1). In mVP24, α5 has different properties (Figure 2a,b and Table 2), while α6 is just a small turn (Figure 1c). These differences in the properties of AHs involved in binding KPNA in eVP24 to those in mVP24 strongly indicates that mVP24 is not immunosuppressive, as is widely accepted18 (or at least it does not use the same mechanism).
Table 1. Residues in Ebola Zaire VP24 (ezVP24, PDBid:4U2XA) that make contact with human karyopherin (PDBid:4U2XD).
One or more atoms from these residues are within 4 Å of residues from human karyopherin.
Table 2. Properties of α5 in VP24 proteins from different strains of Ebola and Marburg.
It can be seen that the Marburg VP24 (mVP24) protein has a distinctly different charge residue composition in the helix. This strongly indicates that mVP24 might not bind human karyopherin, which is the mechanism of immunosuppression by the Ebola VP24 proteins. HM: Hydrophobic moment, RPNR: Ratio of the positive to the negative residues, Len: length of the helix, NCH: number of charged residues.
PDB.Helix
Description
Len
HM
RPNR
NCH
4M0QA.α5
Ebola Zaire Apo
13
2.2
0
1
4U2XA.α5
Ebola Zaire in complex with KPNA
16
4.4
0
2
4D9OA.α5
Ebola Reston Apo
15
3
0
1
3VNEA.α5
Ebola Sudan Apo
14
4.1
0
1
4OR8A.α5
Marburg Apo
16
4.9
0.7
3
Figure 2. Edmundson wheel for α5 of VP24 in ZEBOV strain (eZVP24), Marburg (mVP24) and REBOV (erVP24) viruses.
The color coding for the Edmundson wheel is as follows: all hydrophobic residues are colored red, while hydrophilic residues are colored in blue: dark blue for positively charged residues, medium blue for negatively charged residues and light blue for amides. (a) Apo ezVP24 (PDBid:4M0QA). (b) Apo mVP24 (PDBid:3VNEA). It can be seen that mVP24 has two positively charged residues in the AH, unlike eZVP24. (c) ezVP24 (PDBid:4U2XA) in complex with human karyopherin (PDBid:4U2XD). Note, that Glu113 and Pro114 are now part of the AH, in contrast to the apo AH in (a). (d) Apo erVP24 (PDBid:4D9OA).
S140R substitution in α6 may explain why Ebola Reston strain is non-pathogenic in humans
The REBOV strain ‘does not represent an immediate public health menace on the scale of the African Ebola virus’9, possibly due to the generation of antibodies against this strain39. Also, gene expression study of infected cells showed that the ZEBOV and Marburg viruses has fewer activated IFN-inducible genes relative to REBOV40. Thus, most likely, the REBOV strain does not have the same immunosuppressive capabilities as the ZEBOV or Sudan strain. While α5 of erVP24 has properties similar to ezVP24 (Figure 2c), α6 in REBOV VP24 (erVP24) is clearly different in hydrophobic moment and residue composition (Figure 3). For example, Arg140 in ezVP24 is replaced with Ser140 in erVP24.
Figure 3. Edmundson wheel for α6 of VP24 in ezVP24, esVP24 and erVP24 viruses.
(a) apo ezVP24 (PDBid:4M0QA). (b) ezVP24 in complex with humans karyopherin (PDBid:4U2X). Note, that the AH is extended by two residues (E143 and Q144) as compared to the apo protein. However, the hydrophobic moment remains the same. (c) α6 of esVP24 (PDBid:3VNEA). (d) α6 of erVP24 (PDBid:3VNEA). It can be seen REBOV VP24 has a different hydrophobic moment than the other, since Ser140 is place of Arg140.
To better visualize and quantify this difference, we docked KPNA to erVP24. First, we modelled the apo erVP24 (PDBid:4D9OA) using the ezVP24 complexed with KPNA (PDBid:4U2X) using SWISS-MODEL24. Subsequently, KPNA was docked to this protein using DOCLASP25.
Figure 4 shows ezVP24 and erVP24 docked to KPNA. In ezVP2, KPNA binding is primarily facilitated by electrostatic attraction between the negatively charged Asp124 in α5 and Lys481 in KPNA (at 3.9 Å)12, and a hydrogen bond between Arg140 (α6) and Glu475 of KPNA (among other hydrogen bonds, Table 3). Also, the ezVP24 itself is stabilized by an electrostatic bond between the negatively charged Glu113/OE1 (α5) and the positively charged Arg140/NH1 (α6) at 3.4 Å. Note, that this pair is at distance of 12.8 Å in the apo ezVP24 (PDBid:4M0QA). This 8Å conformational change in these AHs emphasizes the role of plasiticity in binding KPNA. In contrast, in the erVP24, the distance between Glu113/OE1 and Ser140/OG changes from 14 Å in the apo enzyme to 6.2 Å in the docked model. Also, the Ser140/OG atom is not positively charged unlike Arg140/NH1. Further, the possibility of Ser140/OG making a hydrogen bond with Glu475 of KPNA is remote, since they are 6.7 Å apart. Thus, we conclude that the mutation R140S is likely to be one of the critical factors for the non-pathogenic nature of REBOV, since this mutation renders erVP24 incapable of binding KPNA. Other factors might include the different susceptibilities of the glycoproteins of ZEBOV and REBOV for furin cleavage41.
Figure 4. Docking human karyopherin (KPNA) to erVP24.
The erVP24 was modelled using SWISS-MODEL24 using ezVP24 structure complexed with KPNA (PDBid:4U2XA) (See 4D9OA4U2XA.pdb in Dataset 1). The docking was done using DOCLASP25, which superimposes the proteins as well. (a) Superimposition of modelled erVP24 and ezVP24, with bound KPNA. (b) Electrostatic attraction between the negatively charged Glu113/OE1 (α5) and the positively charged Arg140/NH1 (α6) at 3.4 Å, and a hydrogen bond between Arg140 (α6) and Glu475 of KPNA stabilizes the binding. (c) Ser140 replaces Arg140 in erVP24, and fails to make any of the above interactions.
Table 3. Atoms from ZEBOV VP24 (ezVP24) that are closest to the human karyopherin (KPNA) in PDBid:4U2X.
The complete sorted list can be found in ‘4U2XA.4U2XD.maxdist.out.sort’ in Dataset 1. Note, that there is a hydrogen bond between Arg140/NH2 and Glu475/O.
ezVP24 atom
KPNA atom
Distance (Å)
THR/138/OG1
ASP/480/OD2
2.7
ASN/185/ND2
ASP/431/O
2.7
ASN/185/OD1
ARG/398/NH1
2.8
THR/138/N
ASP/480/OD2
2.9
ARG/140/NH2
GLU/475/O
3.0
Docking mouse KPNA to erVP24
We used KPNA from (Mus musculus (mouse) (PDBid:1Y2AC, 50.3% identity and 77.8% similar) to compare the binding of VP24 to KPNA from another related species42. Although, REBOV is pathogenic in non-human primates, there are no known structures for KNPA in other primates (See blastkpna.png in Dataset 1). Figure 5 shows the sequence alignment, the superimposed proteins and the mouse KPNA docked to erVP24 using DOCLASP (See RESTONVP24mouse.p1m in Dataset 1). Note, that the interacting residues (Glu475 and Lys481) are conserved. The fact that erVP24 is not immunosuppressive for mouse is further substantiated by a recent study that noted viral replication in all rodents tested, but disease progression occurs only in STAT1 knockouts43. Note, that erVP24 is able to directly bind STAT1 at levels similar to VP24 from other species21. However, apparently this binding is not sufficient to inhibit the IFN signalling pathway43. Thus, VP24 and its ability to bind KPNA plays a major role in the ‘Reston-pathogenicity puzzle’44. Several putative sites, including a ‘cluster of Reston-specific residues in VP24 is L136, R139 and S140’, have been identified using deuterium exchange mass spectrometry methods44. Our computational method, with its associated caveats, identifies the S140 residue as being more critical than the other sites.
Figure 5. Docking mouse karyopherin (KPNA) to erVP24.
The erVP24 was modelled by SWISS-MODEL24 using ezVP24 structure complexed with KPNA (PDBid:1Y2AC) (See RESTONVP24mouse.p1M in Dataset 1). The docking was done using DOCLASP25, which superimposes the proteins as well. (a) Sequence alignment of human and mouse KPNA, showing that the interacting residues are conserved. (b) Superimposition of human (in cyan) and mouse (in wheat) KPNA done using MUSTANG. (c) Docked mouse KPNA (in wheat) to erVP24 (in limegreen). Interacting residues of mouse KNPA residues (Glu475 and Lys481) make similar contact to erVP24.
Role of intrinsically disordered stretches in VP24
It is interesting to note that the apo α5 (PDBid:4M0QA) is extended by two residues towards the N-terminal (Figure 2c, Glu113 and Pro114) in the ezVP24 complex with KPNA (PDBid:4U2XA). Notably, Pro and Glu are the two most disorder-promoting residues45. The peptide stretch preceding Glu113 in the Sudan Ebola VP24 (PDBid:3VNEA) is also disordered, and residues in that stretch are unassigned in the crystal structure (Figure 1a). Quite interestingly, the α6 (Figure 3a) is also extended by two residues (towards the C-terminal) in the ezVP24 complex (Figure 3d). As mentioned earlier, this stretch is not a helix in mVP24. In the apo Sudan Ebola VP24, α6 (Figure 3c) is similar to the ezVP24 complex (Figure 3b), and is already extended. This is probably due to the fact that Glu is replaced by Asp, which is not disordered-generating. Also, the hydrophobic moment of all three AHs have (almost) the same direction and magnitude (Figure 3a–c). These observations emphasizes the role of intrinsically disordered regions in viral functionality46,47.
Conclusions
The ability of a single mutation to significantly alter the immunosuppressive properties of the Ebola proteins is well established26,27,48. Sequence-based methods (whole genome profiling) are typically used to identify these critical mutations26. Structural studies provide an alternative, and possibly more rational, method to identify such mutations. For example, while double (and not single) mutations are required in VP35 to inhibit protein kinase R activation, it is difficult to rationalize this based on sequence data only28. In the current work, we build on previous work that characterized AH structures in Ebola proteins to rationalize the lack of immunosuppressive properties in the mVP24. ezVP24 binds to KPNA via two AHs (α5 and α5), loops and a residue on a β-sheet. We attribute the lack of immunosuppressive properties of mVP24 to its inability to bind KPNA, which emanates from different characteristics of mVP24 α5 compared to ezVP24 α5. Subsequently, we demonstrate that a single mutation in α6 in the erVP24 might endow it with immunosuppressive properties. We corroborate this conclusion by modelling the apo structure of the erVP24 based on the structure of ezVP24 in complex with KPNA using SWISS-MODEL24, and by docking KPNA to the modelled structure using DOCLASP25. The REBOV strain, first identified in monkeys and imported into the United States from the Philippines8, has never caused disease in humans9,10. However, the isolation of the REBOV strains from pigs in the Philippines29,30, and recently in China31, highlights the significance of finding preventive therapies in the probable scenario that a mutant REBOV for VP24 with immunosuppressive capabilities gets transferred to human handlers. Such a difference does not exist in the VP35 protein, where REBOV VP35 has been used as a model to show how they could silence and sequester double-stranded RNA, which is a key event in immunosuppression49. We also reiterate the potential of using these AHs from VP24 as epitopes50,51 for generating antibodies32,52,53, or innovating drugs to inhibit protein-protein interaction33,54–58. The presence of two intrinsically disordered residues proximal to these AHs in the apo structure that gain a AH structure upon binding should encourage antibody search to use both apo and complexed AHs. It is certainly worth investigating whether supplementing ZMapp, a cocktail of three antibodies that has shown reversion of advanced Ebola symptoms in non-human primates59, with more antibodies would prove more effective.
Data availability
F1000Research: Dataset 1. Version 2. Data used for SCALPEL search methodology to identify plant alpha helical - antimicrobial peptides in the PDB database.
SC wrote the computer programs. All authors analyzed the data, and contributed equally to the writing and subsequent refinement of the manuscript.
Competing interests
No competing interests were disclosed.
Grant information
AMD wishes to acknowledge grant support from the California Department of Food and Agriculture PD/GWSS Board. BJ acknowledges financial support from Tata Institute of Fundamental Research (Department of Atomic Energy). Additionally, BJR is thankful to the Department of Science and Technology for the JC Bose Award Grant. BA acknowledges financial support from the Science Institute of the University of Iceland.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgements
We are grateful to Mary Lou Mendum for her inputs in preparing the manuscript.
Faculty Opinions recommended
References
1.
Dolnik O, Kolesnikova L, Becker S:
Filoviruses: Interactions with the host cell.
Cell Mol Life Sci.
2008; 65(5): 756–776. PubMed Abstract
| Publisher Full Text
2.
Kissling RE, Robinson RQ, Murphy FA, et al.:
Agent of disease contracted from green monkeys.
Science.
1968; 160(3830): 888–890. PubMed Abstract
| Publisher Full Text
3.
Pattyn S, van der Groen G, Courteille G, et al.:
Isolation of Marburg-like virus from a case of haemorrhagic fever in Zaire.
Lancet.
1977; 1(8011): 573–574. PubMed Abstract
| Publisher Full Text
6.
Piot P, Muyembe JJ, Edmunds WJ:
Ebola in west Africa: from disease outbreak to humanitarian crisis.
Lancet Infect Dis.
2014; 14(11): 1034–1035. PubMed Abstract
| Publisher Full Text
7.
Daugherty MD, Malik HS:
How a virus blocks a cellular emergency access lane to the nucleus, STAT!
Cell Host Microbe.
2014; 16(2): 150–152. PubMed Abstract
| Publisher Full Text
8.
Jahrling PB, Geisbert TW, Dalgard DW, et al.:
Preliminary report: isolation of Ebola virus from monkeys imported to USA.
Lancet.
1990; 335(8688): 502–505. PubMed Abstract
| Publisher Full Text
9.
Miranda ME, White ME, Dayrit MM, et al.:
Seroepidemiological study of filovirus related to Ebola in the Philippines.
Lancet.
1991; 337(8738): 425–426. PubMed Abstract
| Publisher Full Text
10.
Miranda ME, Miranda NL:
Reston ebolavirus in humans and animals in the Philippines: a review.
J Infect Dis.
2011; 204(Suppl 3): S757–S760. PubMed Abstract
| Publisher Full Text
12.
Chakraborty S, Rao B, Asgeirsson B, et al.:
Characterizing alpha helical properties of Ebola viral proteins as potential targets for inhibition of alpha-helix mediated protein-protein interactions [v1; ref status: awaiting peer review, http://f1000r.es/4lg].
F1000Res.
2014; 3: 251. Publisher Full Text
13.
Weissenhorn W, Carfi A, Lee KH, et al.:
Crystal structure of the Ebola virus membrane fusion subunit, GP2, from the envelope glycoprotein ectodomain.
Mol cell.
1998; 2(5): 605–616. PubMed Abstract
| Publisher Full Text
14.
Lee JE, Fusco ML, Hessell AJ, et al.:
Structure of the Ebola virus glycoprotein bound to an antibody from a human survivor.
Nature.
2008; 454(7201): 177–182. PubMed Abstract
| Publisher Full Text
| Free Full Text
15.
Xu W, Edwards MR, Borek DM, et al.:
Ebola virus VP24 targets a unique NLS binding site on karyopherin alpha 5 to selectively compete with nuclear import of phosphorylated STAT1.
Cell Host Microbe.
2014; 16(2): 187–200. PubMed Abstract
| Publisher Full Text
| Free Full Text
16.
Suzuki Y, Gojobori T:
The origin and evolution of Ebola and Marburg viruses.
Mol Biol Evol.
1997; 14(8): 800–806. PubMed Abstract
| Publisher Full Text
17.
Feldmann H, Nichol ST, Klenk HD, et al.:
Characterization of filoviruses based on differences in structure and antigenicity of the virion glycoprotein.
Virology.
1994; 199(2): 469–473. PubMed Abstract
| Publisher Full Text
18.
Valmas C, Grosch MN, Schűmann M, et al.:
Marburg virus evades interferon responses by a mechanism distinct from Ebola virus.
PLoS Pathog.
2010; 6(1): e1000721. PubMed Abstract
| Publisher Full Text
| Free Full Text
20.
Reid SP, Leund LW, Hartman AL, et al.:
Ebola virus VP24 binds karyopherin alpha1 and blocks STAT1 nuclear accumulation.
J Virol.
2006; 80(11): 5156–5167. PubMed Abstract
| Publisher Full Text
| Free Full Text
21.
Zhang AP, Bornholdt ZA, Liu T, et al.:
The Ebola virus interferon antagonist VP24 directly binds STAT1 and has a novel, pyramidal fold.
PLoS Pathog.
2012; 8(2): e1002550. PubMed Abstract
| Publisher Full Text
| Free Full Text
22.
Radzimanowski J, Effantin G, Weissenhorn W:
Conformational plasticity of the Ebola virus matrix protein.
Protein Sci.
2014; 23(11): 1519–27. PubMed Abstract
| Publisher Full Text
24.
Arnold K, Bordoli L, Kopp J, et al.:
The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling.
Bioinformatics.
2006; 22(2): 195–201. PubMed Abstract
| Publisher Full Text
25.
Chakraborty S:
DOCLASP - Docking ligands to target proteins using spatial and electrostatic congruence extracted from a known holoenzyme and applying simple geometrical transformations [v1; ref status: awaiting peer review, http://f1000r.es/48g].
F1000Res.
2014; 3: 262. Publisher Full Text
26.
Hartman AL, Ling L, Nichol ST, et al.:
Whole-genome expression profiling reveals that inhibition of host innate immune response pathways by Ebola virus can be reversed by a single amino acid change in the VP35 protein.
J Virol.
2008; 82(11): 5348–5358. PubMed Abstract
| Publisher Full Text
| Free Full Text
27.
Yen B, Mulder LC, Martinez O, et al.:
Molecular Basis for Ebolavirus VP35 Suppression of Human Dendritic Cell Maturation.
J Virol.
2014; 88(21): 12500–12510. PubMed Abstract
| Publisher Full Text
28.
Schűmann M, Gantke T, Műhlberger E:
Ebola virus VP35 antagonizes PKR activity through its C-terminal interferon inhibitory domain.
J Virol.
2009; 83(17): 8993–8997. PubMed Abstract
| Publisher Full Text
| Free Full Text
29.
Marsh GA, Haining J, Robinson R, et al.:
Ebola Reston virus infection of pigs: clinical significance and transmission potential.
J Infect Dis.
2011; 204(Suppl 3): S804–S809. PubMed Abstract
| Publisher Full Text
30.
Barrette RW, Metwally SA, Rowland JM, et al.:
Discovery of swine as a host for the Reston ebolavirus.
Science.
2009; 325(5937): 204–206. PubMed Abstract
| Publisher Full Text
31.
Pan Y, Zhang W, Cui L, et al.:
Reston virus in domestic pigs in China.
Arch Virol.
2014; 159(5): 1129–1132. PubMed Abstract
| Publisher Full Text
32.
Wilson JA, Bray M, Bakken R, et al.:
Vaccine potential of Ebola virus VP24, VP30, VP35, and VP40 proteins.
Virology.
2001; 286(2): 384–390. PubMed Abstract
| Publisher Full Text
33.
Azzarito V, Long K, Murphy NS, et al.:
Inhibition of α-helix-mediated protein-protein interactions using designed molecules.
Nat Chem.
2013; 5(3): 161–173. PubMed Abstract
| Publisher Full Text
34.
Joosten RP, te Beek TA, Krieger E, et al.:
A series of PDB related databases for everyday needs.
Nucleic Acids Res.
2011; 39(Database issue): D411–419. PubMed Abstract
| Publisher Full Text
| Free Full Text
35.
Jones MK, Anantharamaiah GM, Segrest JP:
Computer programs to identify and classify amphipathic alpha helical domains.
J Lipid Res.
1992; 33(2): 287–296. PubMed Abstract
36.
Larkin MA, Blackshields G, Brown NP, et al.:
Clustal W and Clustal X version 2.0.
Bioinformatics.
2007; 23(21): 2947–2948. PubMed Abstract
| Publisher Full Text
37.
Gouy M, Guindon S, Gascuel O:
SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building.
Mol Biol Evol.
2010; 27(2): 221–224. PubMed Abstract
| Publisher Full Text
38.
Konagurthu AS, Whisstock JC, Stuckey PJ, et al.:
MUSTANG: a multiple structural alignment algorithm.
Proteins.
2006; 64(3): 559–574. PubMed Abstract
| Publisher Full Text
39.
Ksiazek TG, West CP, Rollin PE, et al.:
ELISA for the detection of antibodies to Ebola viruses.
J Infect Dis.
1999; 179(Suppl 1): S192–S198. PubMed Abstract
| Publisher Full Text
40.
Kash JC, Műhlberger E, Carter V, et al.:
Global suppression of the host antiviral response by Ebola- and Marburgviruses: increased antagonism of the type I interferon response is associated with enhanced virulence.
J Virol.
2006; 80(6): 3009–3020. PubMed Abstract
| Publisher Full Text
| Free Full Text
41.
Volchkov VE, Feldmann H, Volchkova VA, et al.:
Processing of the Ebola virus glycoprotein by the proprotein convertase furin.
Proc Natl Acad Sci U S A.
1998; 95(10): 5762–5767. PubMed Abstract
| Free Full Text
42.
Kakuk A, Friedländer E, Vereb G Jr, et al.:
Nuclear and nucleolar localization signals and their targeting function in phosphatidylinositol 4-kinase PI4K230.
Exp Cell Res.
2008; 314(13): 2376–2388. PubMed Abstract
| Publisher Full Text
43.
de Wit E, Munster VJ, Metwally SA, et al.:
Assessment of rodents as animal models for Reston ebolavirus.
J Infect Dis.
2011; 204(Suppl 3): S968–S972. PubMed Abstract
| Publisher Full Text
| Free Full Text
45.
Uversky VN:
The alphabet of intrinsic disorder. II. Various roles of glutamic acid in ordered and intrinsically disordered proteins.
Intrinsically Disord Proteins.
2013; 1(1): 18–40. Publisher Full Text
49.
Kimberlin CR, Bornholdt ZA, Li S, et al.:
Ebolavirus VP35 uses a bimodal strategy to bind dsRNA for innate immune suppression.
Proc Natl Acad Sci U S A.
2010; 107(1): 314–319. PubMed Abstract
| Publisher Full Text
| Free Full Text
50.
Takada A, Feldmann H, Stroeher U, et al.:
Identification of protective epitopes on Ebola virus glycoprotein at the single amino acid level by using recombinant vesicular stomatitis viruses.
J Virol.
2003; 77(2): 1069–1074. PubMed Abstract
| Publisher Full Text
| Free Full Text
51.
Wilson JA, Hevey M, Bakken R, et al.:
Epitopes involved in antibody-mediated protection from Ebola virus.
Science.
2000; 287(5458): 1664–1666. PubMed Abstract
| Publisher Full Text
52.
Takada A, Ebihara H, Jones S, et al.:
Protective efficacy of neutralizing antibodies against Ebola virus infection.
Vaccine.
2007; 25(6): 993–999. PubMed Abstract
| Publisher Full Text
53.
Qiu X, Alimonti JB, Melito PL, et al.:
Characterization of Zaire Ebolavirus glycoprotein-specific monoclonal antibodies.
Clin Immunol.
2011; 141(2): 218–227. PubMed Abstract
| Publisher Full Text
54.
Wells JA, McClendon CL:
Reaching for high-hanging fruit in drug discovery at protein-protein interfaces.
Nature.
2007; 450(7172): 1001–1009. PubMed Abstract
| Publisher Full Text
55.
Chapman RN, Dimartino G, Arora PS:
A highly stable short alpha-helix constrained by a main-chain hydrogen-bond surrogate.
J Am Chem Soc.
2004; 126(39): 12252–12253. PubMed Abstract
| Publisher Full Text
56.
Bird GH, Madani N, Perry AF, et al.:
Hydrocarbon double-stapling remedies the proteolytic instability of a lengthy peptide therapeutic.
Proc Natl Acad Sci U S A.
2010; 107(32): 14093–14098. PubMed Abstract
| Publisher Full Text
| Free Full Text
57.
Bird GH, Boyapalle S, Wong T, et al.:
Mucosal delivery of a double-stapled RSV peptide prevents nasopulmonary infection.
J Clin Invest.
2014; 124(5): 2113–24. PubMed Abstract
| Publisher Full Text
| Free Full Text
58.
Harrison RS, Shepherd NE, Hoang HN, et al.:
Downsizing human, bacterial, and viral proteins to short water-stable alpha helices that maintain biological potency.
Proc Natl Acad Sci U S A.
2010; 107(26): 11686–11691. PubMed Abstract
| Publisher Full Text
| Free Full Text
60.
Chakraborty S:
Dataset 1, Version 2 in “Computational modeling of the binding of VP24 to human karyopherin reveals differences between Ebola and Marburg viruses that may correlate to their immune suppression and pathogenicity mechanisms”.
F1000Research.
2014. Data Source
1
Plant Sciences Department, University of California, Davis, CA 95616, USA 2
Department of Biological Sciences, Tata Institute of Fundamental Research, Homi Bhabha Road, Mumbai, 400 005, India 3
Science Institute, Department of Biochemistry, University of Iceland, Dunhaga 3, IS-107 Reykjavik, Iceland
AMD wishes to acknowledge grant support from the California Department of Food and Agriculture PD/GWSS Board. BJ acknowledges financial support from Tata Institute of Fundamental Research (Department of Atomic Energy). Additionally, BJR is thankful to the Department of Science and Technology for the JC Bose Award Grant. BA acknowledges financial support from the Science Institute of the University of Iceland.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Chakraborty S, Rao BJ, Asgeirsson B and Dandekar AM. Correlating the ability of VP24 protein from Ebola and Marburg viruses to bind human karyopherin to their immune suppression mechanism and pathogenicity using computational methods [version 2; peer review: 2 approved with reservations]. F1000Research 2014, 3:265 (https://doi.org/10.12688/f1000research.5666.2)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
Zeng Q. Reviewer Report For: Correlating the ability of VP24 protein from Ebola and Marburg viruses to bind human karyopherin to their immune suppression mechanism and pathogenicity using computational methods [version 2; peer review: 2 approved with reservations]. F1000Research 2014, 3:265 (https://doi.org/10.5256/f1000research.6328.r7280)
Zoonotic transmission of Ebola virus (EBOV) to humans causes a severe haemorrhagic fever in affected humans. Neither vaccines nor therapeutics are available at present. To devise antiviral strategies, it is important to understand the pathogenicity and molecular basis of EBOV
... Continue reading
Zoonotic transmission of Ebola virus (EBOV) to humans causes a severe haemorrhagic fever in affected humans. Neither vaccines nor therapeutics are available at present. To devise antiviral strategies, it is important to understand the pathogenicity and molecular basis of EBOV infection. Among all the 7 proteins including NP, VP35, VP40, GP, VP30, VP24 and L of EBOV, structural proteins VP24 and VP35 have already been found playing a key role in interference with proper functioning of host interferon system. Present computational analysis offered insights into potentially underlying mechanisms of VP24.
Suggestions for revision:
The title seems too long. Single-point mutation in VP24 ---one of the key molecular mechanisms underlying the pathogenicity of filovirus.
The writing needs to be substantially improved. There are grammar errors, illogical expression, inaccurate, undefined and misleading descriptions, and some biased or questionable conclusions. Just take the abstract as an example where questionable words by the authors were marked in bold and my opinion labeled in italics.
“Immune response suppression is crucial for viral invasion. The protein VP24 is pivotal in achieving this in Ebolaalthough interestingly the mechanism of immune suppression is different in the closely related Marburg virus[Here there is no error, just that the “Marburg virus” came out suddenly. Maybe the authors want to say: The protein VP24 is pivotal in achieving this in both Ebola and the closely related Marburg virus, although interestingly the mechanism of immune suppression is different]. Here, we illustrate [Illustrated? How? Computationally or experimentally? That is important for a comprehensive and clear understanding of this article] that a possible molecular basis[if it’s really just a “possible basis”, what’s the value to publish a “possible” thing? And what’s the value of your computational analysis?May be should be “one of the key molecular basis”] for this difference emanates from two alpha helical structures (α5 and α6) in VP24 involved in binding human karyopherin (KPNA) (PDBid:4U2X), wherein the Ebola and Marburg viruses have distinctly different charged properties in α5. α6 is absent in Marburg, and [here, the subject is missing. Who/what “has a different hydrophobic moment”? α5 or α6 or something else?]has a different hydrophobic moment in the Reston Ebola (REBOV) species, which is surprisingly [what makes “non-pathogenic in humans” so surprising? The authors know why, and I know why, because we know background information related to Ebola, but the point is, “you know and I know” doesn’t necessarily mean all the readers know why. The background information should be clearly presented with the least words. In all the 5 Ebola species, outbreaks of ZEBOV, SEBOV, CIEBOV and BEBOV have been recorded. However, REBOV has just been detected in swine]non-pathogenic in humans[the only one non-pathogenic in humans out of 5 Ebola species including ZEBOV, SEBOV, CIEBOV and BEBOV. This information actually should be given in the beginning of this abstract]. Based on the hypothesis that REBOV is not immunosuppressive, which is in turn is [here are two “is”? what does “ in turn” mean?] due to its inability to bind KPNA, we show [showed] by docking KPNA[which species? Human, mice or pigs?] to the REBOV VP24 that the single amino acid substitution R140S [what does “R140S” mean? substitution of R to S at 140 residue?] is responsible for this difference between REBOV and Zaire Ebola strains [just based on the analysis of only one protein VP24, could we safely conclude that “is responsible for this difference between REBOV and Zaire Ebola”? Do we have 100% confidence that there is absolutely no other mechanism within VP24 and provided by other proteins, say VP35? This just presents your direct analyzed results including what properties changed, including electrical charge, hydrophobic or hydrophilic, and their binding property to STAT1]. Such a scenario of getting a virulent REBOV through a single mutation is particularly worrisome, since the REBOV, once found only in monkeys, has been recently detected in pigs. We also reiterate the potential of using these helices as potential epitopes for generating protective antibodies against Ebola.”
The abstract should be logically organized, starting from background information to methods, direct analyzed results, conclusion and finally the significance of present research.
In the introduction, some key information is missing which is indispensable for clear, accurate and logical understanding of the following analysis, related discussion and correlation between analysis and observed facts. For example:
There are Five EBOV species that have been defined, Zaire ebolavirus (ZEBOV), Sudan ebolavirus (SEBOV), Coˆte d’Ivoire ebolavirus (CIEBOV), Bundibugyo ebolavirus (BEBOV) and Reston ebolavirus (REBOV). They have shown different pathogenicity up to date. Outbreaks of ZEBOV, SEBOV, CIEBOV and BEBOV have been recorded. However, REBOV has just been detected in swine.
“In Ebola, the crucial role of host immune system evasion is accomplished by two proteins: VP35 and VP24.” ---What about Marburg? It’s also dependent on VP35 and VP24 or just on VP24? because we are going to compare between Ebola and Marburg.
The main part of the article - computer modeling and analysis of VP24 and its interactions to other molecules - is reliable and sufficient. However, what makes the present analysis valuable is whether these analysis explain observed facts including pathogenicity between Marburg and Ebola virus, and among different Ebola species, and what about experimental findings by others? In other words, are there any experimental observations supporting present analysis?
In the Conclusion part of this article, the authors did not actually conclude their main analyzed results and corresponding significance. This “Conclusion” is actually a discussion.
About the discussion:
As both VP35 and VP24 contribute to “immune evasion” as described in “Introduction”, how could you get an accurate and reliable conclusion just based on the analysis of VP24? Change your angle of view.
All previous experimental observations and conclusions by other scientists about VP24 should be included in discussion, giving a comprehensive and impartial comparative analysis. However, some key studies are obviously missing in this part. For example:
The IFN system can protect immune-competent mice from lethal EBOV infection. Adaptation of ZEBOV to lethal infection of mice was associated with mutations in VP24 and NP (Ebihara et al., 2006). However, both wild-type VP24 and VP24 of the mouse-adapted (MA) strain were able to bind to human and mouse NP-1 importins and to disrupt the interaction with PY-STAT1 (Reid et al., 2007). Similar findings were documented for VP24 of REBOV, which is believed to be non-pathogenic for humans, and it was shown that ZEBOV, REBOV and MA VP24 can suppress IFN-b-induced gene expression (Reid et al., 2007). Thus, alterations in VP24 interference with the IFN response might not account for the acquisition of virulence of MA ZEBOV in mice and for the lack of virulence of REBOV in humans, respectively.
These findings are opposites of the present statement. However, the different, even opposite opinions on the same topic by different scientists are normal phenomenon in scientific community. The most important thing is how to analyze, to explain these differences and finally get a scientific conclusion and evaluation of you own work, without ignoring those opposite findings or opinions.
Competing Interests: No competing interests were disclosed.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Zeng Q. Reviewer Report For: Correlating the ability of VP24 protein from Ebola and Marburg viruses to bind human karyopherin to their immune suppression mechanism and pathogenicity using computational methods [version 2; peer review: 2 approved with reservations]. F1000Research 2014, 3:265 (https://doi.org/10.5256/f1000research.6328.r7280)
McIntosh M. Reviewer Report For: Correlating the ability of VP24 protein from Ebola and Marburg viruses to bind human karyopherin to their immune suppression mechanism and pathogenicity using computational methods [version 2; peer review: 2 approved with reservations]. F1000Research 2014, 3:265 (https://doi.org/10.5256/f1000research.6328.r7131)
That all EBOV species employ VP24 to subvert the host innate immune response by binding KPNA.
That a single point mutation R140S can explain a lack of pathogenesis by REBOV in humans through a lost capacity to bind
... Continue reading
There are two assumptions.
That all EBOV species employ VP24 to subvert the host innate immune response by binding KPNA.
That a single point mutation R140S can explain a lack of pathogenesis by REBOV in humans through a lost capacity to bind KPNA.
Neither of these assumptions have been experimentally verified by the authors or elsewhere. At a minimum, it seems these assumptions would need to be addressed in silico through analysis of the KPNA for a susceptible host to REBOV.
Minor concerns:
Abstract
"which is surprisingly non-pathogenic" ...to .."which is notably non-pathogenic in humans."
"which is in turn is due to its inability to bind" ..to.."which is in turn due to its inability to bind ."
Results and Discussion:
Dataset 1 title and legend appear to be mislabeled as "search methodology to identify plant alpha helica-antimicrobial peptides in the PDB dataset"...should this not be labeled as."to identify filovirus VP24 alpha helices"?
Major Concerns:
Regarding Figure 1 and Figure 4:
Like REBOV, Bundibugyo (BEBOV)and Tai Forest virus (TFV) also have substitutions at the R140 of ZEBOV. BEBOV and TFV have His and Gln in this position instead of the Arg of ZEBOV and SEBOV. As these are both pathogenic in humans, albeit perhaps less so, how might these substitutions compare to putative binding with KPNA?
The added experiment of docking mouse KPNA to erVP24 is appreciated but does not address the important question of whether or not non-human primate KPNA has compensatory substitutions to restore the potential for binding Reston VP24. Following this line of thought, such compensatory substitutions would conversely not be expected to reduce binding with VP24 from other African species of EBOV. While the ability of single point mutations to abrogate protein-protein interactions is indeed well established, the ability of compensatory substitutions to restore intermolecular interactions is also well established. Would it not be more prudent to sequence KPNA from a non-human primate host susceptible to hemorrhagic disease caused by REBOV and test the hypothesis in silico?
Understandably, access to non-human primate sequence is limiting making it difficult to address this concern. In light of the inability to validate these findings either experimentally or in silico with a susceptible host species for fatal disease with REBOV, I suggest that the observation of the R140S subsititution in REBOV and its forecast impact on pathogenicity, while intriguing, remains highly speculative.
Competing Interests: No competing interests were disclosed.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
McIntosh M. Reviewer Report For: Correlating the ability of VP24 protein from Ebola and Marburg viruses to bind human karyopherin to their immune suppression mechanism and pathogenicity using computational methods [version 2; peer review: 2 approved with reservations]. F1000Research 2014, 3:265 (https://doi.org/10.5256/f1000research.6328.r7131)
McIntosh M. Reviewer Report For: Correlating the ability of VP24 protein from Ebola and Marburg viruses to bind human karyopherin to their immune suppression mechanism and pathogenicity using computational methods [version 2; peer review: 2 approved with reservations]. F1000Research 2014, 3:265 (https://doi.org/10.5256/f1000research.6051.r6641)
This article presents an interesting in silico observation to possibly explain observed differences in pathogenesis and suppression of host immune antiviral type 1 interferon (INF) responses emanating from structural differences in VP24 proteins of various Ebola virus (EBOV) species and
... Continue reading
This article presents an interesting in silico observation to possibly explain observed differences in pathogenesis and suppression of host immune antiviral type 1 interferon (INF) responses emanating from structural differences in VP24 proteins of various Ebola virus (EBOV) species and Marburg virus. For context, host antiviral INF signaling is known to induce nuclear transport of tyrosine-phosphorylated signal transducer and activator of transcription 1 (STAT1) as an early stage in a signaling cascade that activates expression of host genes involved in antiviral mechanisms. A subset of the host Karyopherin alpha (KPNA) family are involved in the nuclear transport of activated STAT1, and EBOV VP24 protein has been shown by others (Xu et al., 2014) to bind KPNA thus interfering with this nuclear transport and the progression of host innate and adaptive immune responses to EBOV infection. Marburg virus is noted to interfere with host antiviral INF responses differently via direct inhibition of phosphorylation/activation of STAT1 and STAT2. In this article, in addition to gross charge and structural differences in two alpha helices (a5 and a6) of VP24 between EBOV and Marburg viruses, possibly explaining the different mechanisms of INF response suppression, the authors hypothesis that a single substitution R140S in VP24 between the pathogenic Zaire ebolavirus (ZEBOV) and non-pathogenic Reston ebolavirus (REBOV) alters charged properties of the a5 alpha helix leading to a lack of binding to human KPNA by REBOV VP24. This substitution in REBOV VP24 is hypothesized to be responsible for the lack of REBOV pathogenesis in humans. The authors further express concern regarding the potential for a single amino acid substitution in REBOV, previously observed in domestic swine, to perhaps lead to a more pathogenic virus in the future.
Article Content:
The study employs computational modeling of the primary VP24 amino acid sequences of different EBOV species and Marburg virus onto the previously resolved crystal structure of ZEBOV VP24 bound to KPNA5 (Xu et al., 2014). The direct comparisons between potential binding sites of KPNA and VP24 from different species of EBOV are intriguing but the study unfortunately lacks experimental verification either through in vitro binding or functional studies In addition there are concerns regarding the accuracy of theoretical modeling of primary VP24 sequences from various EBOV species to the known crystal structure of ZEBOV VP24 and KPNA5 peptides. Without experimental verification it is not possible to draw the conclusion that the R140S substitution present in REBOV affects binding to KPNA or that it is responsible for the absence of pathogenicity in humans. One approach not tried is modeling of a KPNA5 homolog from non-human primates as REBOV is known to still be pathogenic in non-human primates. In concept, it seems unlikely that a single mutation could be wholly responsible for the observed differences in pathogenicity between REBOV and other EBOV species. Various mechanisms not involving VP24 including EBOV glycoprotein and VP35-mediated mechanisms of immune suppression as well as a potential host genetic differences are likely to have critical influences on EBOV pathogenesis beyond the specific mechanism of VP24-mediated suppression of activated STAT1 nuclear localization and expression of INF triggered host antiviral mechanisms.
Of minor importance, invasion should be replaced with pathogenesis in the first sentence of the abstract and minor typographical errors should be corrected.
Competing Interests: No competing interests were disclosed.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
McIntosh M. Reviewer Report For: Correlating the ability of VP24 protein from Ebola and Marburg viruses to bind human karyopherin to their immune suppression mechanism and pathogenicity using computational methods [version 2; peer review: 2 approved with reservations]. F1000Research 2014, 3:265 (https://doi.org/10.5256/f1000research.6051.r6641)
Sandeep Chakraborty, Tata Institute of Fundamental Research, India
01 Dec 2014
Author Response
Dear Dr McIntosh,
'We would like to thank you for taking the time to review this paper, and for your insightful comments. While our method is computational, and there is no
...
Continue readingDear Dr McIntosh,
'We would like to thank you for taking the time to review this paper, and for your insightful comments. While our method is computational, and there is no easy way to get around that fact for us with respect to Ebola, we do believe that dissemination of such information can provide direction in the effort to understand, and finally abrogate, the mechanism of pathogenesis of the Ebola virus. Recently, we have used the PAGAL [ref-1] software to design anti-microbial peptides that work against plant pathogens [ref-2].
The logical thread of our hypothesis in this manuscript follows the inability of the VP24 from Marburg to bind KPNA owing to the difference in two helices (analzyed using PAGAL) that bind KPNA in the Zaire Ebola virus. We believe this point is irrefutable. A small difference in one of the helices (alpha6) in the VP24 from Reston Ebola virus results in two computationally arrived differences.
Different hydrophobic moment in the Edmundson wheel (Fig3) (on a known structure, so confirmed). This difference is also visible in a multiple sequence alignment of the protein from different species.
Different charged interactions of the residues in KPNA and VP24, after docking (on a modelled structure, possible inaccuracies).
These differences might not have drawn attention, if Reston Ebola was not known to be non-pathogenic to humans. We have taken care at each point to clearly indicate that this is a possibility, and not a foregone conclusion. In fact, studying the ‘Reston-pathogenicity puzzle’ using deuterium exchange mass spectrometry (DEMS) methods, Zhang et. al. (2012) have identified putative sites which includes a ‘cluster of Reston-specific residues in VP24 is L136, R139 and S140’ [ref-3]. It is possible that these differences would not lead to loss of binding when such experiments are finally done, and we would have to revise our hypothesis (which the F1000Research model allows us to do). We emphasize on the role of computational methods to make intelligent and informed decisions, enabling biologist to design experiments, and minimizing human effort and cost - something that has been sorely missing in the Ebola effort.
In this context, and also in response to your comment on the unlikelihood of a single mutation resulting in pathogenicity, we would like to cite recent work that identifies two mutations (one in VP24 and the other in the nucleoprotein) resulting in the acquisition of high virulence in mice [ref-4]. The VP24 mutation is Thr50, and lies on a beta-sheet, and its importance in the structure has not been completely understood to date, although this residue is another putative site in the DEMS study [ref-3]. Our group, that has focused on the importance of alpha-helices, but not beta-sheets [ref-5], is trying to rationalize the overwhelming significance of this mutation.
We also appreciate your idea of using KPNA from a non-human primate. However, only mice and rats have solved KPNAs. We have now included data on docking of a mouse KPNA to the Reston VP24 after conducting a similar analysis, and found no difference in their interactions (Fig. 5). Interestingly, we have also come across a study which concludes that only a STAT1 knockout mouse is susceptible to Reston Ebola virus [ref-6]. This strongly points towards the lack of immunosuppressive properties of the Reston Ebola virus in mice.
We have also made the suggested minor corrections, and had the manuscript corrected for typographical errors (Mary Mendum has been acknowledged). We hope that we have addressed your concerns by the changes that we have made.
Thanking you,
Sincerely,
Sandeep Chakraborty (Corresponding author)
[References]
[[1|type=journal|title=PAGAL - Properties and corresponding graphics of alpha helical structures in proteins [v2; ref status: indexed, http://f1000r.es/4e7]|authors=Chakraborty/S;Rao/B;Dandekar/A|source=F1000Research|year=2014|vol=3|issue=206|pmid=25352981|doi=10.12688/f1000research.4952.2|url=http://f1000research.com/articles/3-206/v2]] [[2|type=journal|title=The PDB database is a rich source of alpha-helical anti-microbial peptides to combat disease causing pathogens [v1; ref status: awaiting peer review, http://f1000r.es/4sa]|authors=Chakraborty/S;Phu/M;Rao/B;Asgeirsson/B;Dandekar/A|source=F1000Research|year=2014|vol=3|issue=295|doi=10.12688/f1000research.5802.1|url=http://f1000research.com/articles/3-295/v1]] [[3|type=journal|title=The ebolavirus VP24 interferon antagonist: know your enemy|authors=Zhang/AP;Abelson/DM;Bornholdt/ZA;Liu/T;Woods/VL Jr;Saphire/EO|source=Virulence|year=2012|vol=3|issue=5|fpage=440|lpage=445|pmid=23076242|pmcid=3485981|doi=10.4161/viru.21302|url=http://www.landesbioscience.com/journals/viru/abstract.php?id=21302]] [[4|type=journal|title=Molecular determinants of Ebola virus virulence in mice|authors=Ebihara/H;Takada/A;Kobasa/D;Jones/S;Neumann/G;Theriault/S;Bray/M;Feldmann/H;Kawaoka/Y|source=PLoS Pathog|year=2006|vol=2|issue=7|fpage=e73|pmid=16848640|pmcid=1513261 |doi=10.1371/journal.ppat.0020073|url=http://www.plospathogens.org/article/info%3Adoi%2F10.1371%2Fjournal.ppat.0020073]] [[5|type=journal|title=Characterizing alpha helical properties of Ebola viral proteins as potential targets for inhibition of alpha-helix mediated protein-protein interactions [v2; ref status: approved with reservations 1, http://f1000r.es/4qr]|authors=Chakraborty/S;Rao/B;Asgeirsson/B;Dandekar/A|source=F1000Research|year=2014|vol=3|issue=251|doi=10.12688/f1000research.5573.2|url=http://f1000research.com/articles/3-251/v2]] [[6|type=journal|title=Assessment of rodents as animal models for Reston ebolavirus|authors=de Wit/E;Munster/VJ;Metwally/SA;Feldman/H|source=J Infect Dis|year=2011|vol=204|issue=Suppl 3|fpage=S968|lpage=S972|pmid=21987777|pmcid= 3189989|doi=10.1093/infdis/jir330|url=http://jid.oxfordjournals.org/content/204/suppl_3/S968.long]]
Dear Dr McIntosh,
'We would like to thank you for taking the time to review this paper, and for your insightful comments. While our method is computational, and there is no easy way to get around that fact for us with respect to Ebola, we do believe that dissemination of such information can provide direction in the effort to understand, and finally abrogate, the mechanism of pathogenesis of the Ebola virus. Recently, we have used the PAGAL 1 software to design anti-microbial peptides that work against plant pathogens 2.
The logical thread of our hypothesis in this manuscript follows the inability of the VP24 from Marburg to bind KPNA owing to the difference in two helices (analzyed using PAGAL) that bind KPNA in the Zaire Ebola virus. We believe this point is irrefutable. A small difference in one of the helices (alpha6) in the VP24 from Reston Ebola virus results in two computationally arrived differences.
Different hydrophobic moment in the Edmundson wheel (Fig3) (on a known structure, so confirmed). This difference is also visible in a multiple sequence alignment of the protein from different species.
Different charged interactions of the residues in KPNA and VP24, after docking (on a modelled structure, possible inaccuracies).
These differences might not have drawn attention, if Reston Ebola was not known to be non-pathogenic to humans. We have taken care at each point to clearly indicate that this is a possibility, and not a foregone conclusion. In fact, studying the ‘Reston-pathogenicity puzzle’ using deuterium exchange mass spectrometry (DEMS) methods, Zhang et. al. (2012) have identified putative sites which includes a ‘cluster of Reston-specific residues in VP24 is L136, R139 and S140’ 3. It is possible that these differences would not lead to loss of binding when such experiments are finally done, and we would have to revise our hypothesis (which the F1000Research model allows us to do). We emphasize on the role of computational methods to make intelligent and informed decisions, enabling biologist to design experiments, and minimizing human effort and cost - something that has been sorely missing in the Ebola effort.
In this context, and also in response to your comment on the unlikelihood of a single mutation resulting in pathogenicity, we would like to cite recent work that identifies two mutations (one in VP24 and the other in the nucleoprotein) resulting in the acquisition of high virulence in mice 4. The VP24 mutation is Thr50, and lies on a beta-sheet, and its importance in the structure has not been completely understood to date, although this residue is another putative site in the DEMS study 3. Our group, that has focused on the importance of alpha-helices, but not beta-sheets 5, is trying to rationalize the overwhelming significance of this mutation.
We also appreciate your idea of using KPNA from a non-human primate. However, only mice and rats have solved KPNAs. We have now included data on docking of a mouse KPNA to the Reston VP24 after conducting a similar analysis, and found no difference in their interactions (Fig. 5). Interestingly, we have also come across a study which concludes that only a STAT1 knockout mouse is susceptible to Reston Ebola virus 6. This strongly points towards the lack of immunosuppressive properties of the Reston Ebola virus in mice.
We have also made the suggested minor corrections, and had the manuscript corrected for typographical errors (Mary Mendum has been acknowledged). We hope that we have addressed your concerns by the changes that we have made.
Thanking you,
Sincerely,
Sandeep Chakraborty (Corresponding author)
References 1. Chakraborty S, Rao B, Dandekar A: PAGAL - Properties and corresponding graphics of alpha helical structures in proteins [v2; ref status: indexed, http://f1000r.es/4e7]. F1000Research. 2014; 3 (206). PubMed Abstract | Publisher Full Text | Reference Source 2. Chakraborty S, Phu M, Rao B, Asgeirsson B, et al.: The PDB database is a rich source of alpha-helical anti-microbial peptides to combat disease causing pathogens [v1; ref status: awaiting peer review, http://f1000r.es/4sa]. F1000Research. 2014; 3 (295). Publisher Full Text | Reference Source 3. Zhang AP, Abelson DM, Bornholdt ZA, Liu T, et al.: The ebolavirus VP24 interferon antagonist: know your enemy. Virulence. 2012; 3 (5): 440-445 PubMed Abstract | Free Full Text | Publisher Full Text | Reference Source 4. Ebihara H, Takada A, Kobasa D, Jones S, et al.: Molecular determinants of Ebola virus virulence in mice. PLoS Pathog. 2006; 2 (7): e73 PubMed Abstract | Free Full Text | Publisher Full Text | Reference Source 5. Chakraborty S, Rao B, Asgeirsson B, Dandekar A: Characterizing alpha helical properties of Ebola viral proteins as potential targets for inhibition of alpha-helix mediated protein-protein interactions [v2; ref status: approved with reservations 1, http://f1000r.es/4qr]. F1000Research. 2014; 3 (251). Publisher Full Text | Reference Source 6. de Wit E, Munster VJ, Metwally SA, Feldman H: Assessment of rodents as animal models for Reston ebolavirus. J Infect Dis. 2011; 204 (Suppl 3): S968-S972 PubMed Abstract | Free Full Text | Publisher Full Text | Reference Source
Competing Interests:No competing interests were disclosed.Close
Sandeep Chakraborty, Tata Institute of Fundamental Research, India
01 Dec 2014
Author Response
Dear Dr McIntosh,
'We would like to thank you for taking the time to review this paper, and for your insightful comments. While our method is computational, and there is no
...
Continue readingDear Dr McIntosh,
'We would like to thank you for taking the time to review this paper, and for your insightful comments. While our method is computational, and there is no easy way to get around that fact for us with respect to Ebola, we do believe that dissemination of such information can provide direction in the effort to understand, and finally abrogate, the mechanism of pathogenesis of the Ebola virus. Recently, we have used the PAGAL [ref-1] software to design anti-microbial peptides that work against plant pathogens [ref-2].
The logical thread of our hypothesis in this manuscript follows the inability of the VP24 from Marburg to bind KPNA owing to the difference in two helices (analzyed using PAGAL) that bind KPNA in the Zaire Ebola virus. We believe this point is irrefutable. A small difference in one of the helices (alpha6) in the VP24 from Reston Ebola virus results in two computationally arrived differences.
Different hydrophobic moment in the Edmundson wheel (Fig3) (on a known structure, so confirmed). This difference is also visible in a multiple sequence alignment of the protein from different species.
Different charged interactions of the residues in KPNA and VP24, after docking (on a modelled structure, possible inaccuracies).
These differences might not have drawn attention, if Reston Ebola was not known to be non-pathogenic to humans. We have taken care at each point to clearly indicate that this is a possibility, and not a foregone conclusion. In fact, studying the ‘Reston-pathogenicity puzzle’ using deuterium exchange mass spectrometry (DEMS) methods, Zhang et. al. (2012) have identified putative sites which includes a ‘cluster of Reston-specific residues in VP24 is L136, R139 and S140’ [ref-3]. It is possible that these differences would not lead to loss of binding when such experiments are finally done, and we would have to revise our hypothesis (which the F1000Research model allows us to do). We emphasize on the role of computational methods to make intelligent and informed decisions, enabling biologist to design experiments, and minimizing human effort and cost - something that has been sorely missing in the Ebola effort.
In this context, and also in response to your comment on the unlikelihood of a single mutation resulting in pathogenicity, we would like to cite recent work that identifies two mutations (one in VP24 and the other in the nucleoprotein) resulting in the acquisition of high virulence in mice [ref-4]. The VP24 mutation is Thr50, and lies on a beta-sheet, and its importance in the structure has not been completely understood to date, although this residue is another putative site in the DEMS study [ref-3]. Our group, that has focused on the importance of alpha-helices, but not beta-sheets [ref-5], is trying to rationalize the overwhelming significance of this mutation.
We also appreciate your idea of using KPNA from a non-human primate. However, only mice and rats have solved KPNAs. We have now included data on docking of a mouse KPNA to the Reston VP24 after conducting a similar analysis, and found no difference in their interactions (Fig. 5). Interestingly, we have also come across a study which concludes that only a STAT1 knockout mouse is susceptible to Reston Ebola virus [ref-6]. This strongly points towards the lack of immunosuppressive properties of the Reston Ebola virus in mice.
We have also made the suggested minor corrections, and had the manuscript corrected for typographical errors (Mary Mendum has been acknowledged). We hope that we have addressed your concerns by the changes that we have made.
Thanking you,
Sincerely,
Sandeep Chakraborty (Corresponding author)
[References]
[[1|type=journal|title=PAGAL - Properties and corresponding graphics of alpha helical structures in proteins [v2; ref status: indexed, http://f1000r.es/4e7]|authors=Chakraborty/S;Rao/B;Dandekar/A|source=F1000Research|year=2014|vol=3|issue=206|pmid=25352981|doi=10.12688/f1000research.4952.2|url=http://f1000research.com/articles/3-206/v2]] [[2|type=journal|title=The PDB database is a rich source of alpha-helical anti-microbial peptides to combat disease causing pathogens [v1; ref status: awaiting peer review, http://f1000r.es/4sa]|authors=Chakraborty/S;Phu/M;Rao/B;Asgeirsson/B;Dandekar/A|source=F1000Research|year=2014|vol=3|issue=295|doi=10.12688/f1000research.5802.1|url=http://f1000research.com/articles/3-295/v1]] [[3|type=journal|title=The ebolavirus VP24 interferon antagonist: know your enemy|authors=Zhang/AP;Abelson/DM;Bornholdt/ZA;Liu/T;Woods/VL Jr;Saphire/EO|source=Virulence|year=2012|vol=3|issue=5|fpage=440|lpage=445|pmid=23076242|pmcid=3485981|doi=10.4161/viru.21302|url=http://www.landesbioscience.com/journals/viru/abstract.php?id=21302]] [[4|type=journal|title=Molecular determinants of Ebola virus virulence in mice|authors=Ebihara/H;Takada/A;Kobasa/D;Jones/S;Neumann/G;Theriault/S;Bray/M;Feldmann/H;Kawaoka/Y|source=PLoS Pathog|year=2006|vol=2|issue=7|fpage=e73|pmid=16848640|pmcid=1513261 |doi=10.1371/journal.ppat.0020073|url=http://www.plospathogens.org/article/info%3Adoi%2F10.1371%2Fjournal.ppat.0020073]] [[5|type=journal|title=Characterizing alpha helical properties of Ebola viral proteins as potential targets for inhibition of alpha-helix mediated protein-protein interactions [v2; ref status: approved with reservations 1, http://f1000r.es/4qr]|authors=Chakraborty/S;Rao/B;Asgeirsson/B;Dandekar/A|source=F1000Research|year=2014|vol=3|issue=251|doi=10.12688/f1000research.5573.2|url=http://f1000research.com/articles/3-251/v2]] [[6|type=journal|title=Assessment of rodents as animal models for Reston ebolavirus|authors=de Wit/E;Munster/VJ;Metwally/SA;Feldman/H|source=J Infect Dis|year=2011|vol=204|issue=Suppl 3|fpage=S968|lpage=S972|pmid=21987777|pmcid= 3189989|doi=10.1093/infdis/jir330|url=http://jid.oxfordjournals.org/content/204/suppl_3/S968.long]]
Dear Dr McIntosh,
'We would like to thank you for taking the time to review this paper, and for your insightful comments. While our method is computational, and there is no easy way to get around that fact for us with respect to Ebola, we do believe that dissemination of such information can provide direction in the effort to understand, and finally abrogate, the mechanism of pathogenesis of the Ebola virus. Recently, we have used the PAGAL 1 software to design anti-microbial peptides that work against plant pathogens 2.
The logical thread of our hypothesis in this manuscript follows the inability of the VP24 from Marburg to bind KPNA owing to the difference in two helices (analzyed using PAGAL) that bind KPNA in the Zaire Ebola virus. We believe this point is irrefutable. A small difference in one of the helices (alpha6) in the VP24 from Reston Ebola virus results in two computationally arrived differences.
Different hydrophobic moment in the Edmundson wheel (Fig3) (on a known structure, so confirmed). This difference is also visible in a multiple sequence alignment of the protein from different species.
Different charged interactions of the residues in KPNA and VP24, after docking (on a modelled structure, possible inaccuracies).
These differences might not have drawn attention, if Reston Ebola was not known to be non-pathogenic to humans. We have taken care at each point to clearly indicate that this is a possibility, and not a foregone conclusion. In fact, studying the ‘Reston-pathogenicity puzzle’ using deuterium exchange mass spectrometry (DEMS) methods, Zhang et. al. (2012) have identified putative sites which includes a ‘cluster of Reston-specific residues in VP24 is L136, R139 and S140’ 3. It is possible that these differences would not lead to loss of binding when such experiments are finally done, and we would have to revise our hypothesis (which the F1000Research model allows us to do). We emphasize on the role of computational methods to make intelligent and informed decisions, enabling biologist to design experiments, and minimizing human effort and cost - something that has been sorely missing in the Ebola effort.
In this context, and also in response to your comment on the unlikelihood of a single mutation resulting in pathogenicity, we would like to cite recent work that identifies two mutations (one in VP24 and the other in the nucleoprotein) resulting in the acquisition of high virulence in mice 4. The VP24 mutation is Thr50, and lies on a beta-sheet, and its importance in the structure has not been completely understood to date, although this residue is another putative site in the DEMS study 3. Our group, that has focused on the importance of alpha-helices, but not beta-sheets 5, is trying to rationalize the overwhelming significance of this mutation.
We also appreciate your idea of using KPNA from a non-human primate. However, only mice and rats have solved KPNAs. We have now included data on docking of a mouse KPNA to the Reston VP24 after conducting a similar analysis, and found no difference in their interactions (Fig. 5). Interestingly, we have also come across a study which concludes that only a STAT1 knockout mouse is susceptible to Reston Ebola virus 6. This strongly points towards the lack of immunosuppressive properties of the Reston Ebola virus in mice.
We have also made the suggested minor corrections, and had the manuscript corrected for typographical errors (Mary Mendum has been acknowledged). We hope that we have addressed your concerns by the changes that we have made.
Thanking you,
Sincerely,
Sandeep Chakraborty (Corresponding author)
References 1. Chakraborty S, Rao B, Dandekar A: PAGAL - Properties and corresponding graphics of alpha helical structures in proteins [v2; ref status: indexed, http://f1000r.es/4e7]. F1000Research. 2014; 3 (206). PubMed Abstract | Publisher Full Text | Reference Source 2. Chakraborty S, Phu M, Rao B, Asgeirsson B, et al.: The PDB database is a rich source of alpha-helical anti-microbial peptides to combat disease causing pathogens [v1; ref status: awaiting peer review, http://f1000r.es/4sa]. F1000Research. 2014; 3 (295). Publisher Full Text | Reference Source 3. Zhang AP, Abelson DM, Bornholdt ZA, Liu T, et al.: The ebolavirus VP24 interferon antagonist: know your enemy. Virulence. 2012; 3 (5): 440-445 PubMed Abstract | Free Full Text | Publisher Full Text | Reference Source 4. Ebihara H, Takada A, Kobasa D, Jones S, et al.: Molecular determinants of Ebola virus virulence in mice. PLoS Pathog. 2006; 2 (7): e73 PubMed Abstract | Free Full Text | Publisher Full Text | Reference Source 5. Chakraborty S, Rao B, Asgeirsson B, Dandekar A: Characterizing alpha helical properties of Ebola viral proteins as potential targets for inhibition of alpha-helix mediated protein-protein interactions [v2; ref status: approved with reservations 1, http://f1000r.es/4qr]. F1000Research. 2014; 3 (251). Publisher Full Text | Reference Source 6. de Wit E, Munster VJ, Metwally SA, Feldman H: Assessment of rodents as animal models for Reston ebolavirus. J Infect Dis. 2011; 204 (Suppl 3): S968-S972 PubMed Abstract | Free Full Text | Publisher Full Text | Reference Source
Competing Interests:No competing interests were disclosed.Close
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Spreadsheet data files may not format correctly if your computer is using different default delimiters (symbols used to separate values into separate cells) - a spreadsheet created in one region is sometimes misinterpreted by computers in other regions. You can change the regional settings on your computer so that the spreadsheet can be interpreted correctly.
How to fix it
Save downloaded CSV file
Open spreadsheet program (e.g. Excel)
Click the ‘Data’ tab at the top
Click the ‘From text’ icon (top left)
Browse for downloaded CSV file, click ‘Import’
Ensure ‘Delimited’ radio button is selected, click ‘Next’
Check one of the appropriate delimiter checkboxes (you can visualize the formatting by looking at the data preview below these options)
Chakraborty S, Rao BJ, Asgeirsson B and Dandekar AM. Dataset 1 in: Correlating the ability of VP24 protein from Ebola and Marburg viruses to bind human karyopherin to their immune suppression mechanism and pathogenicity using computational methods. F1000Research 2014, 3:265 (https://doi.org/10.5256/f1000research.5666.d40354)
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
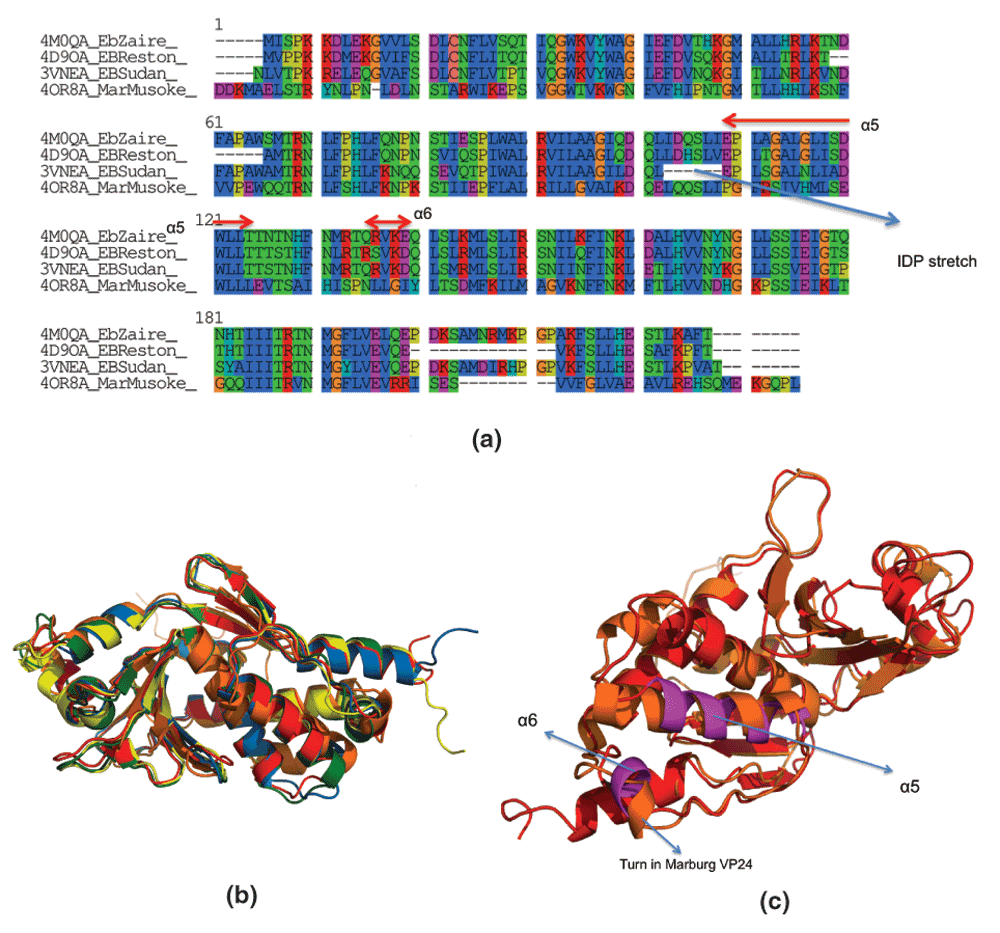
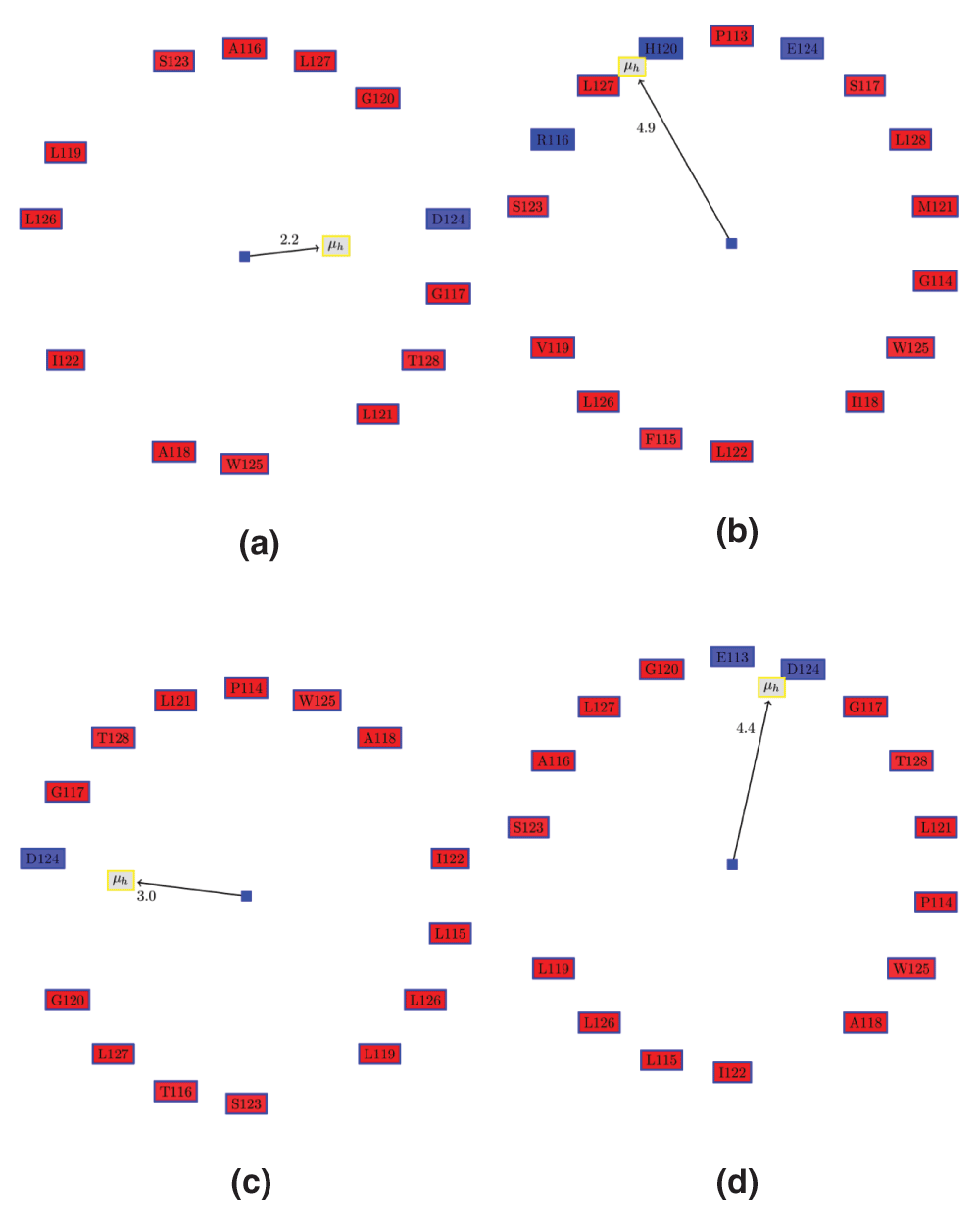
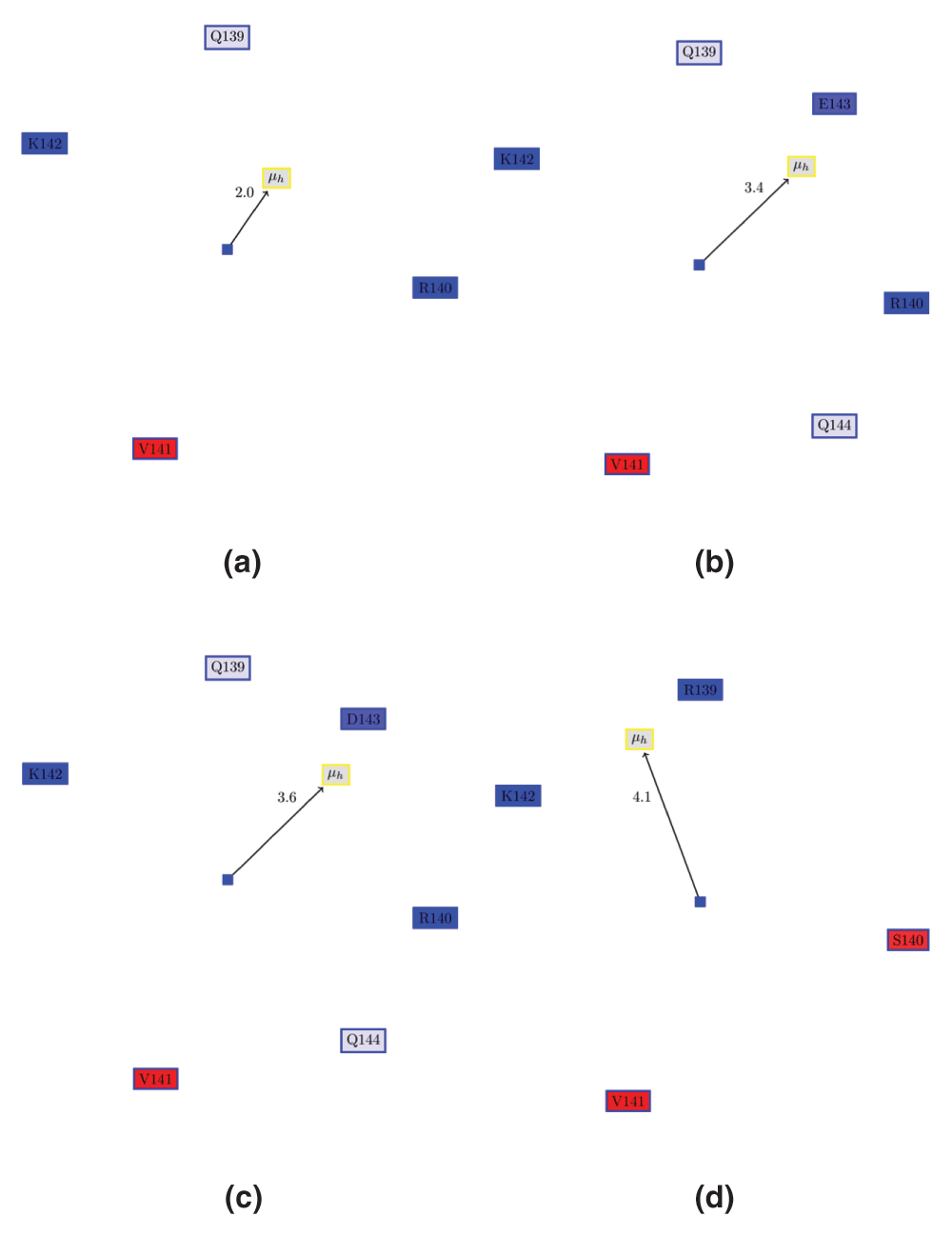
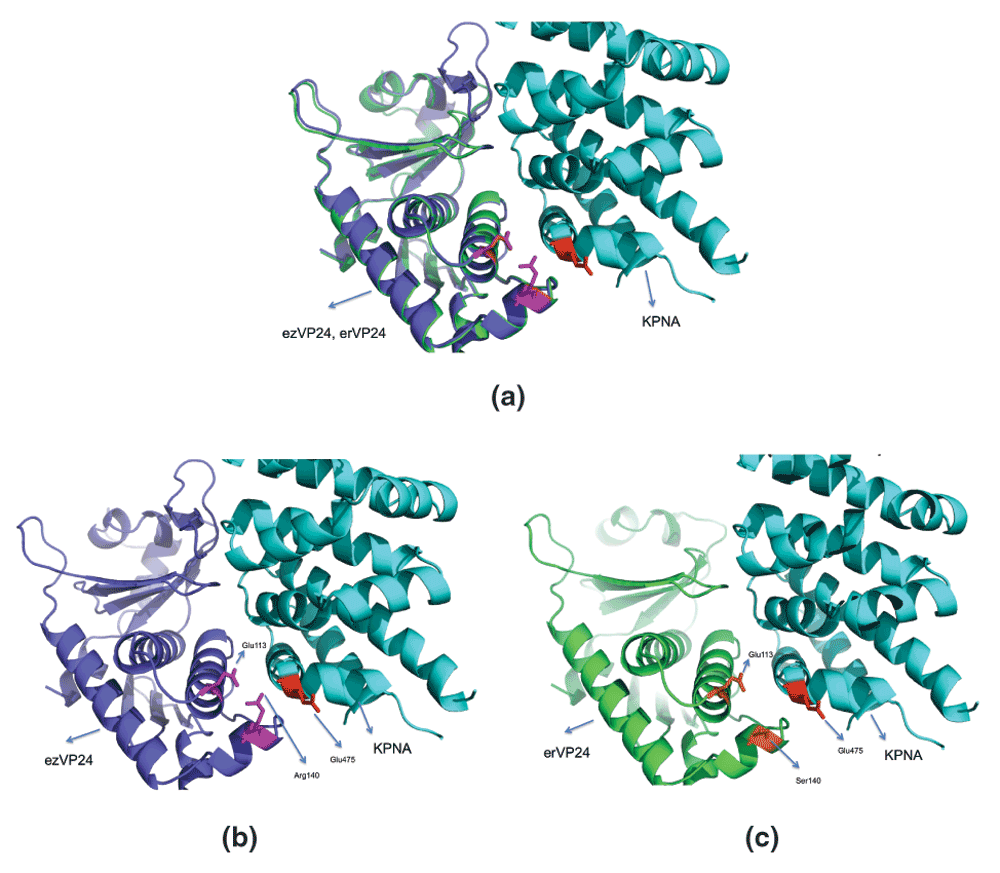
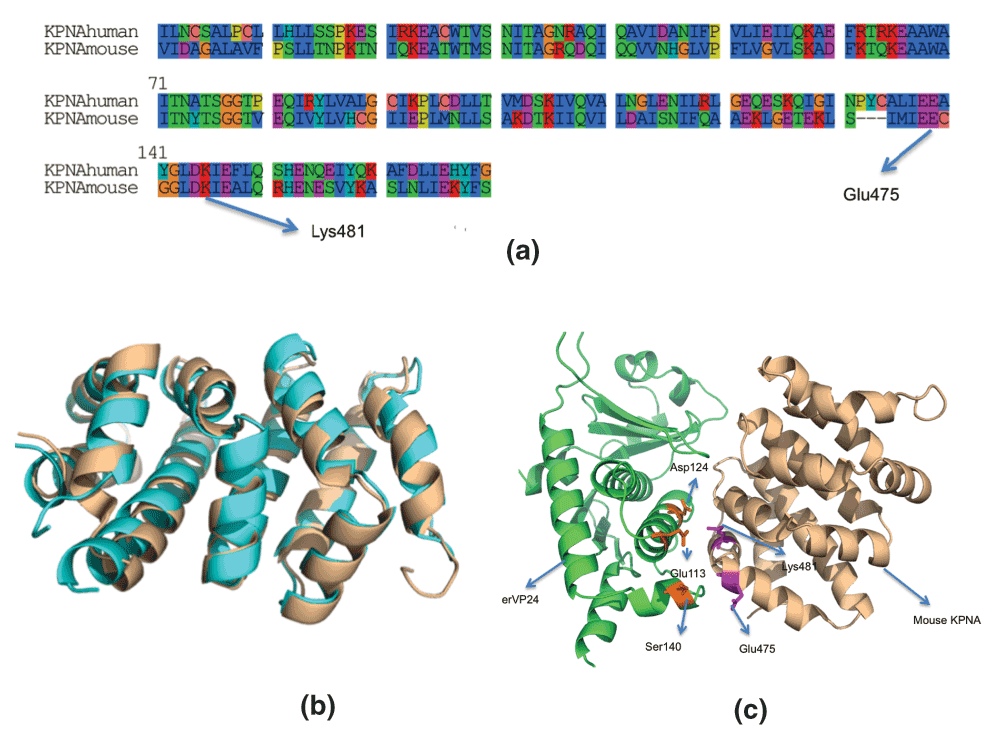
Comments on this article Comments (0)