Keywords
Antimicrobials, Biofilms, E. coli, Environmental
Antimicrobials, Biofilms, E. coli, Environmental
This rewritten version, more focused on the blue Gram stain itself, contains a less speculative discussion regarding the origins and implications of cell clumping.
See the author's detailed response to the review by Klaus Kayser
See the author's detailed response to the review by Venkataramana Kandi
See the author's detailed response to the review by William R. Jacobs
The virtual colony count (VCC) microbiological assay has been used for over a decade to measure the effect of antimicrobial peptides such as defensins and LL-37 against a variety of bacteria. (Ericksen et al., 2005; Zhao et al., 2013). It infers antimicrobial activity based on the quantitative growth kinetics of 200 μL batch cultures of bacteria grown in 96-well plates using a method of enumeration of viable cells (Brewster, 2003) mathematically identical to the method of enumeration of amplicons utilized by quantitative real-time PCR (Heid et al., 1996). The originally published plate configuration included a ring of 36 wells containing uninoculated Mueller Hinton Broth (MHB) capable of detecting cross-contamination (Ericksen, 2014). There was evidence that bacteria might form clumps and biofilms during the assay, including scatter detectable by the plate reader and the presence of ubiquitous macroscopic clumping in tryptic soy broth. 10 μL samples of cross-contamination control wells that had become turbid after VCC experiments were Gram-stained, revealing few clumps. Apparently, most clumps were not retained on the glass during the Gram stain procedure (Gram, 1884), whether fixed to the slide by heat or methanol. The application of lactophenol cotton blue, ordinarily used to visualize fungi by staining cell wall polysaccharides such as chitin, revealed circles and rings consistent with the caramelized residue of polysaccharides, which presumably included capsular polysaccharides and slime secreted concomitantly with clump and biofilm formation. These indigo circles and rings could be consistent either with a heterogeneous subpopulation of E. coli or with slight contamination with a second strain.
The VCC assay was conducted using the 36 edge wells to detect contamination as originally described (Ericksen et al., 2005), except a rectangular piece of Parafilm M (6 × 0.25 squares) was wrapped around the 96-well plate before the start of the 2-hour and 12-hour plate reader runs. Parafilm strips remained almost entirely intact and in place throughout the 12-hour run at 37°C and resulted in the complete absence of dust large enough to be visible using an Olympus 8Z61 crystallographic microscope on the ledge between the 96 wells and the edge of the plate, except for a single speck in one experiment observed near a crack in the Parafilm. Parafilm also prevented the visible decrease in edge well volume due to evaporation that originally necessitated excluding these wells from the experimental portion of the assay (Ericksen et al., 2005). 10 µL samples of edge wells were added to droplets of sterile water or media and spread on Mueller Hinton Agar, Tryptic Soy Agar, and Sabouraud’s Agar plates. Colonies were analyzed by morphology, wet mounts, Gram stains, and biochemical analysis using Becton Dickinson Enteropluri Product Number 261185.
Glass slides were scrubbed with PCMX hand soap using a pipe cleaner. 10 µL of cells sampled from 96-well plates after VCC assays using twice-concentrated MHB in the outgrowth step were added to the slides and equilibrated to ambient humidity overnight. The slides were heat-fixed by placing the sample at the point in space at the upper tip of the inner blue flame of a Bunsen burner three times for one second each, removing the slide for one second in between (Figure 1C). Ambient relative humidity was 40–60%. The slides were stained with Fluka Analytical Gram Staining Kit Product Number 77730 and again equilibrated to ambient humidity overnight in a vertical position. Becton Dickinson Lactophenol Cotton Blue Stain Droppers Product Number 261188 were applied to the Gram stained sample and digital images were captured using an Amscope light microscope at 160×, 400× and 1600× magnification and Toupview software. The Adobe Photoshop thresholding function was applied to the 400× digital images using a threshold of 100. Black pixels were enumerated using the histogram function.
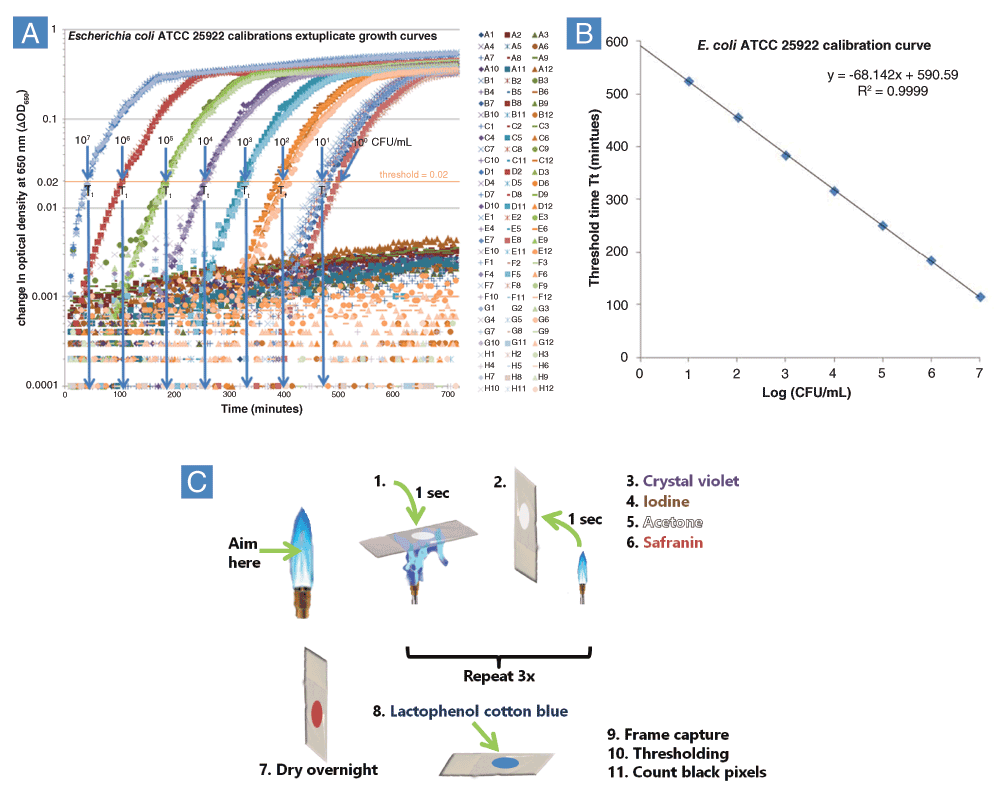
A: Escherichia coli ATCC 25922 was diluted 10-fold in Mueller-Hinton Broth and growth kinetics was monitored for 12h using a Tecan infinite M1000 plate reader according to the UMB calibration procedure (Ericksen et al., 2005). QGK was used to calculate threshold times, Tt. Scatter below the threshold of 0.02 suggested cell clumping. B: Calibration curve relating Tt to the logarithm of cell concentration in a separate experiment. Both experiments shown in A and B were conducted in November, 2011 by Le Zhao. C: Lactophenol Cotton Blue Gram Stain Procedure. Overnight steps allowed for equilibration to the ambient humidity during summer months in the IHV building at UMB, which ranged from 40–60%. Water content and temperature may be important factors for the caramelization process to be quantitatively reproducible.
Macroscopic clumps were observed in 25 mL TSB batch cultures of E. coli ATCC® 25922™ grown at 37°C in early exponential phase to an expected optical density at 650 nm (OD650) of approximately 0.3. A 1 mL uncovered sample placed in a cuvette and cooled to room temperature rapidly formed small macroscopic clumps (up to about 1 mm in diameter), some of which exhibited motility, swimming in a synchronized wave downward to form a single large macroscopic clump (up to 1 cm long, equal to the cuvette width) at the base of the cuvette. OD650 plummeted up to 2% per minute, reaching equilibrium after a 10–20% decrease when placed in a room temperature HPLC detector, as cells in suspension joined the clump beneath the light path. The optical density readings declined so rapidly that only the first two digits of the four reported by the Waters 600 detector could be recorded. Observing cuvettes containing such clumps, it was apparent that cohesion, rather than adhesion, was more important, since the clumps moved downward from one corner to the other corner of the cuvette as it was rotated by hand.
Macroscopic clumping in the batch culture or cuvette outside the detector was no longer observed after four changes: 1. using a small HEPA-filtered air purifier, 2. replacing in-house deionized Milli-Q water with purchased molecular biology grade water, 3. replacing 2×MHB prepared and autoclaved in-house using reusable jars with Teknova 2× cation-adjusted MHB, and 4. filter-sterilizing phosphate buffers made near the portable air purifier, rather than autoclaving in reusable jars. Even after these remediation measures, uncovered 1 mL samples placed in the detector for 2 hours formed a macroscopic clump at the base of the cuvette accompanied by a decrease in optical density, suggesting that at least one clumping environmental factor (CEF) was concentrated by the fan and filter within detector acting as a dust trap. Thus, 1 mL samples of E. coli ATCC® 25922™ served as biosensors for CEFs, and the detector served as a biosensor positive control. Corner-seeking motility of E. coli ATCC® 25922™ was also observed on MH agar plates wrapped in Parafilm and incubated at room temperature for 2–3 weeks, as indicated by the formation of a ~1 cm-wide confluent ring around the entire edge of the plate, even though confluent areas and single colonies that originally appeared after 1–2 days were separate from the edge. The UMB VCC procedure was sensitive to cross-contamination in the 36 uninoculated edge wells, possibly indicating that clumping affects the particle size distribution and adhesive properties of the cells, which in turn promotes aerosol formation during pipetting (Ericksen, 2014). Figure 2 depicts cells sampled from a cross-contaminated edge well after storage at 4°C. The UCLA VCC method, with cells in 10 µL pipetted beneath 90 µL rather than a 50 µL suspension added to 50 µL as droplets from above, (Welkos et al., 2011) minimizes the probability of cross-contamination and is a safer and more effective method of transferring bacteria such as the hazardous BSL-3 pathogen Bacillus anthracis.
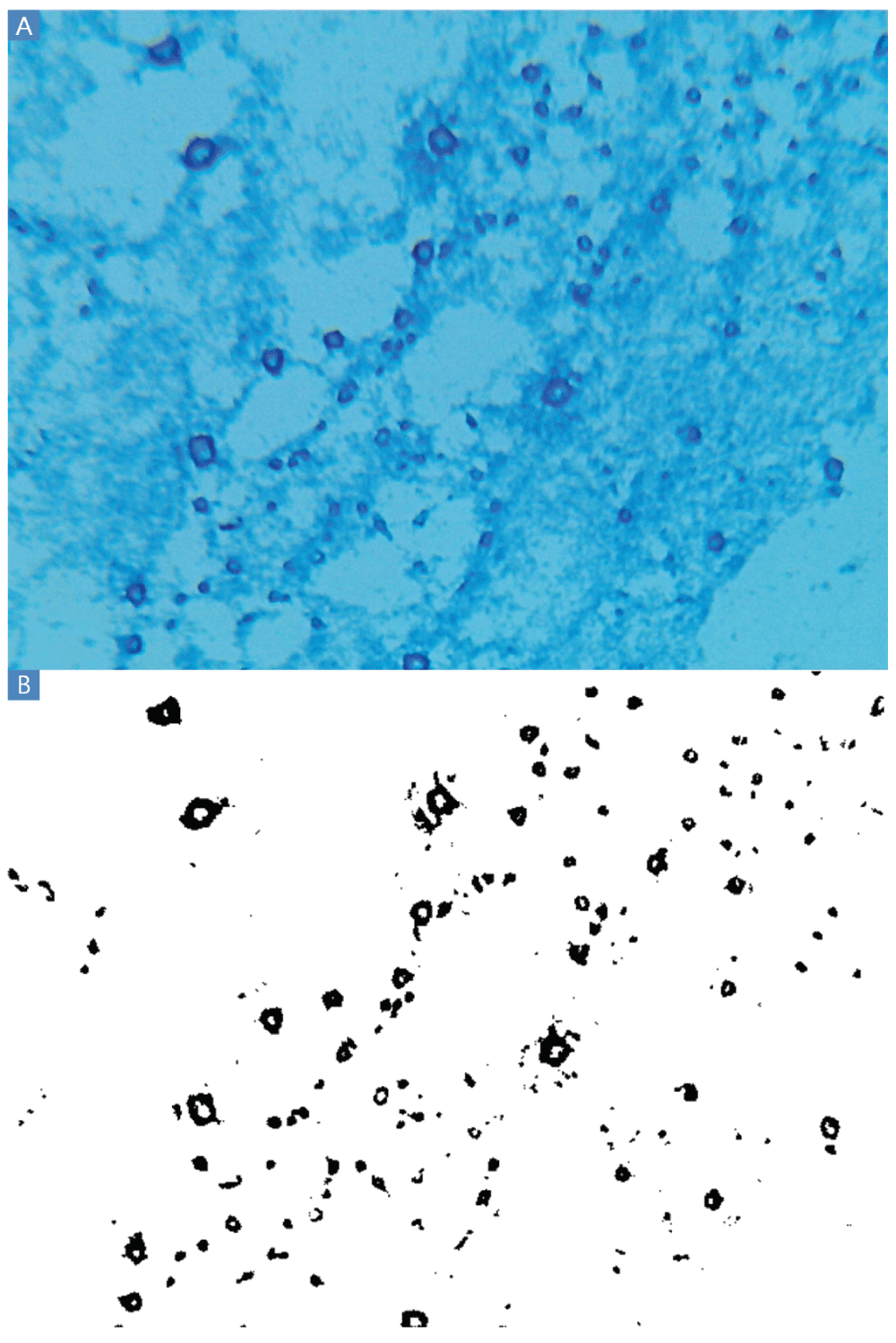
A: Blue rings indicate the polysaccharide residue of clumps of cells presumably washed from the slides during the Gram stain procedure. These polysaccharides were invisible when inspected after Gram staining and before application of lactophenol cotton blue. Other experiments produced smaller dark blue circles rather than rings. B: Thresholding results. A large majority of black pixels are contained within the polysaccharide rings. The VCC experiments resulting in cross-contamination and Escherichia coli ATCC 25922 clumps with dark blue BGS staining were conducted by author BE in the summer of 2013 at UMB.
The lactophenol cotton blue Gram stain (BGS) revealed ubiquitous circular or ring-shaped structures that stained dark blue (Figure 2A). All cells stained light blue because all cells are glycosylated and concentrate polysaccharides from the media as part of their metabolism. Rare regions of indistinct blue staining were also observed, probably resulting from starch and other polysaccharides present in MHB, suggesting that the intensity of blue staining could also arise from starch and other carbohydrates with the capsular polysaccharides. MHB contains 1.5 g/L starch, plus a variety of other carbohydrates contained in beef extract. Carbohydrates, which must have included Maillard reaction (Maillard, 1912) and caramelization products, adhered to the glass in the intense heat of the fixation steps and endured on the slide throughout the Gram stain procedure. These polysaccharide residues had been invisible when these same slides were observed after Gram staining and before application of lactophenol cotton blue. The intensity of dark blue staining suggests copious capsule and slime formation.
Applying the thresholding technique using a threshold of 100 differentiated the dark from the light staining with little apparent background noise (Figure 2B). Thresholding of BGS images captured at 160× and 1600× magnification (Figure 3) are also possible using the Amscope microscope. However, pixelation could add imprecision at 160× and the large size of clumps would increase variability from field to field at 1600×. TSB or MHB cultures of E. coli ATCC® 43827™ (ML-35) produced no macroscopic clumps under any conditions in several experiments conducted in 2013 and 2014, indicating that the influence of CEFs is strain-dependent.
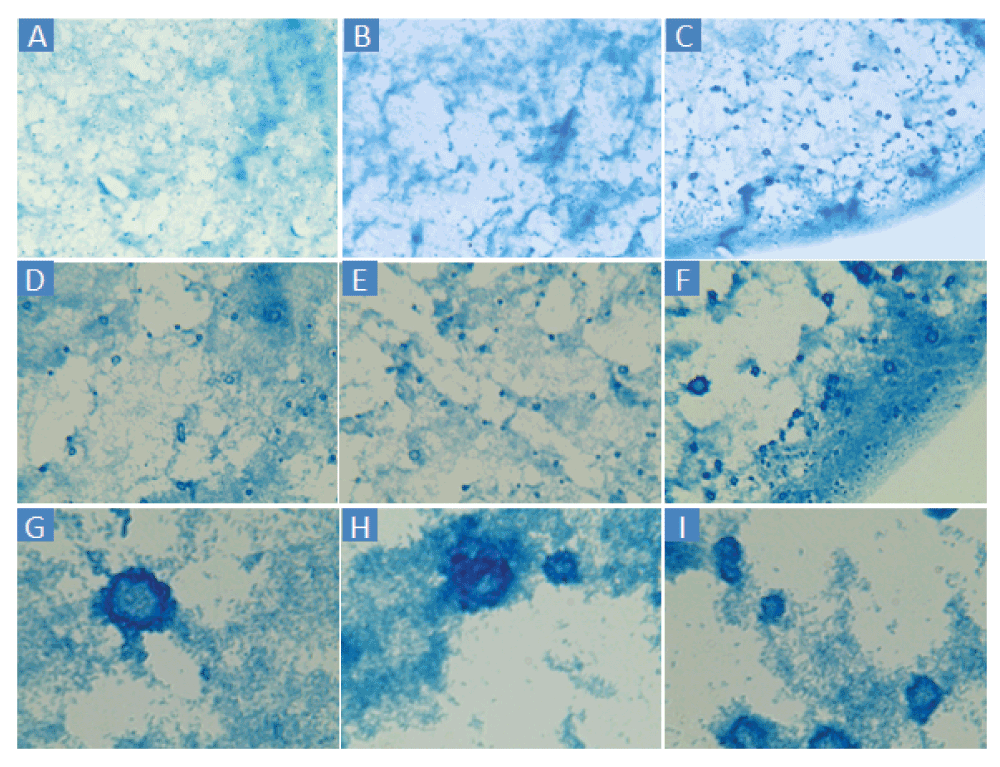
A–C: 160×. D–F: 400×. G–I: 1600×. Cells were sampled from the edge wells of a different virtual colony count experiment than Figure 2.
The history of hundreds of VCC experiments at UMB between 2003 and 2014 clearly shows that edge wells are almost always clear, not turbid, after the 12h outgrowth phase of VCC experiments. In a 1-month period in August and September 2013, I conducted 13 quadruplicate calibration experiments using the same pipetting technique as the sextuplicate calibration experiments in the original VCC publication (Ericksen et al., 2005). However, in the 2013 experiments, four, rather than six, calibration curves were confined to 32 internal wells (C3-F10). These experiments used the rich media MHB, TSB or slight variations thereof. The external 64 wells (rows A, B, G and H and columns 1, 2, 11 and 12) contained two rings of contamination control wells rather than the single ring of 36 wells originally used. In these experiments conducted just outside a biosafety cabinet used for VCC experiments, none of the 832 contamination control wells turned turbid after the 12h incubation. These experiments strongly suggest that CEFs present in the laboratory environment are overwhelmingly non-culturable in rich media such as MHB or TSB. In addition to viable contamination, unculturable bacteria could exert an influence upon rapidly growing E. coli cells. Furthermore, nucleic acids are known to cause cells to coalesce into clumps over a broad size distribution in both bacterial and mammalian cell culture. Airborne CEFs smaller than a bacterial cell could pass through the HEPA filters with little or no resistance, meaning that these molecules could have affected experiments conducted both inside and outside biosafety cabinets. Measures such as trypsinization, treatment with other proteases, and treatment with nucleases such as benzonase are commonly employed to reduce or eliminate clumping (Kruse & Patterson, 1973). For the same purpose, shear was employed in VCC calibration curves by placing pipette tips in contact with the cross-sectional corner of each well when pipetting up and down 15 times to mix (Ericksen, 2014), although growth curves showed evidence of clumps large enough to produce measurable differences in optical density that preceded exponential growth (Figure 1A, region between a change in optical density of 0.001 and 0.01). Clumping had no effect on the linearity of the calibration curve (Figure 1B), possibly indicating that a fraction of cells routinely grow as clumps and biofilms in the absence of antimicrobial agents.
The presence of polysaccharides associated with E. coli ATCC® 25922™ cohesion suggests that in the conditions studied at UMB, this strain employs clumping as a defense mechanism in response to the presence of one or more CEF. Forming a clump surrounded by polysaccharides could contribute to resistance to antimicrobial lectins such as defensins (Wang et al., 2003) that would be bound and inhibited at the surface, limiting further inward diffusion and protecting persister cells (Ericksen et al., 2005) at the center of the clump. These survivors could contribute to the deviation from simple exponential killing (Luria & Latarjet, 1947) observed throughout all VCC studies at UMB of defensin activity against E. coli. MHB contains a considerable amount (1.5 g/L) of added starch. Polysaccharides in rich media could contribute to the complete inhibition of antimicrobial peptides, which is essential for VCC assays to be capable of enumerating surviving bacteria by the QGK data analysis method. Qualitative defensin lectin activity generally follows the hierarchy: glycosylated proteins > branched polysaccharides > linear polysaccharides > oligosaccharides > monosaccharides. (Lehrer, R. I., personal communication) Bacterial slime and capsules are highly branched and contain glycosylated proteins (Wilkinson, 1958). If inhibition follows the same qualitative pattern as binding, bacterial capsular polysaccharides would be potent defensin inhibitors. Clump, biofilm and capsule formation may have evolved partially as resistance mechanisms to the ancient selection pressure exerted throughout the tree of life by antimicrobial peptides in the environment.
A possible consequence of the inhibition of defensins by polysaccharides could be that therapies with lectin antimicrobial peptides as active ingredients would not be effective against clumps or biofilms in the absence of at least one other active ingredient that degrades the polysaccharide capsule, such as a glycosidase. Because polysaccharide structures in capsules and slime vary widely, as do glycosidase substrate specificities, any given enzyme might be active against only a narrow range of bacteria. In the absence of in vivo glycosidases, activity against a broad spectrum of pathogenic bacteria would therefore require an enzyme cocktail of glycosidases accompanying the lectin antimicrobial peptide or a glycosidase with unusually promiscuous substrate specificity.
figshare: Blue Gram Stain images from three cross-contamination edge wells of a Virtual Colony Count assay at 160×, 400×, or 1600×, doi: http://dx.doi.org/10.6084/m9.figshare.1269193 (Ericksen, 2014).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)