Park S, Jung Y, An SWA et al. RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan [version 4; peer review: 3 approved]. F1000Research 2017, 6:1515 (https://doi.org/10.12688/f1000research.12337.4)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan
[version 4; peer review: 3 approved]
Previously titled: RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan
Sangsoon Park
https://orcid.org/0000-0002-5008-1609
1, Yoonji Jung1, Seon Woo A. An1, [...] Heehwa G. Son1, Wooseon Hwang1, Dongyeop Lee1, Murat Artan1, Hae-Eun H. Park1, Dae-Eun Jeong1, Yujin Lee1, Seung-Jae V. Lee
https://orcid.org/0000-0002-6103-156X
1,2
Sangsoon Park
https://orcid.org/0000-0002-5008-1609
1, Yoonji Jung1, [...] Seon Woo A. An1, Heehwa G. Son1, Wooseon Hwang1, Dongyeop Lee1, Murat Artan1, Hae-Eun H. Park1, Dae-Eun Jeong1, Yujin Lee1, Seung-Jae V. Lee
1Department of Life Sciences, Pohang University of Science and Technology, Pohang, Gyeongbuk, 37673, South Korea 2School of Interdisciplinary Bioscience and Bioengineering, Pohang University of Science and Technology, Pohang, Gyeongbuk, 37673, South Korea
Seung-Jae V. Lee
Roles:
Conceptualization,
Funding Acquisition,
Project Administration,
Supervision,
Validation,
Writing – Original Draft Preparation,
Writing – Review & Editing
OPEN PEER REVIEW
REVIEWER STATUS
Abstract
Background: α-arrestins are a family of proteins that are implicated in multiple biological processes, including metabolism and receptor desensitization. Methods: Here, we sought to examine the roles of α-arrestins in the longevity of Caenorhabditis elegans through an RNA interference screen. Results: We found that feeding worms with bacteria expressing double-stranded RNA against each of 24 out of total 29 C. elegans α-arrestins had little effect on lifespan. Thus, individual C. elegans α-arrestins may have minor effects on longevity. Conclusions: This study will provide useful information for future research on the functional role of α-arrestins in aging and longevity.
Keywords
C. elegans, α-arrestin, insulin/IGF-1 signaling, aging, lifespan
Corresponding author:
Seung-Jae V. Lee
Competing interests:
No competing interests were disclosed.
Grant information:
This work was supported by the Korean Government (MSIP) through the National Research Foundation of Korea (NRF) [NRF-2016R1E1A1A01941152] funded to S-J.V.L.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
α-arrestins are a family of proteins that contain arrestin domains whose sequences and structures have similarities with those of classical visual and β-arrestins1–3. α-arrestins are considered as ancestral forms of arrestins because their orthologs exist in fungi, including yeast, which do not have visual or β-arrestins1,4. Several mammalian α-arrestins modulate metabolism and receptor desensitization5,6, but much remains to be elucidated concerning the functions of α-arrestins in many organisms. The human genome encodes 6 α-arrestins and 4 visual or β-arrestins. Interestingly, the simple roundworm Caenorhabditis elegans genome contains 29 α-arrestin and 1 β-arrestin genes1,7. Therefore, the C. elegans system provides opportunities for the genetic analysis of α-arrestins in various aspects of physiology both individually and combinatorially. However, information regarding the functions of C. elegans α-arrestins is limited8,9.
C. elegans is an excellent genetic model organism that has been exploited for studying conserved biological processes, including apoptosis, behavior, development and aging. In particular, its short lifespan in combination with genetic amenability has made C. elegans one of the most popular models for research on aging and longevity10,11. Many factors, including components in insulin/insulin-like growth factor 1 (IGF-1) signaling (IIS), have been identified as lifespan and aging regulators in C. elegans11–13. For example, genetic inhibition of IIS components, such as DAF-2/insulin/IGF-1 receptor, robustly extends lifespan and delays physiological aging through up-regulating transcription factors, including DAF-16/FOXO11–13. Importantly, the roles of these aging-regulatory factors in C. elegans have been shown to be conserved in other species, including Drosophila and mammals12,13. One of the powerful ways to identify novel factors that influence aging is by employing genetic screens, such as an RNA interference (RNAi) screen. We previously identified several genetic factors, including RNA helicases, that modulate longevity in C. elegans, through targeted or genome-wide RNAi screens14–17. Because of its robust longevity phenotype that confers sensitivity to changes in lifespan, daf-2/insulin/IGF-1 receptor mutants serve as an excellent platform for the identification of novel lifespan-regulating factors17,18.
In this study, we aimed to determine whether any α-arrestins played a role in the lifespan regulation of wild-type or daf-2 mutants. We performed a lifespan assay-based RNAi screen targeting 24 out of 29 C. elegans α-arrestins. We found that α-arrestin double-stranded RNA (dsRNA)-expressing E. coli feeding had little effect on the lifespan of wild-type or daf-2 mutants. Thus, C. elegans α-arrestins may play minor or modulatory roles in lifespan regulation. Based on our results, it will be important to test the roles of α-arrestins in combinatorial manners and/or by using strong loss-of-function mutations in future research.
Methods
Caenorhabditis elegans strains
All strains were maintained as previously described19. The following strains were used in this study: N2 wild-type, CF1041 daf-2(e1370) III outcrossed six times to N2.
Phylogenetic analysis
The protein sequences of 27 α-arrestins, except arrd (arrestin domain protein)-20 and arrd-21, were obtained from Wormbase (www.wormbase.org, version WS259). The protein sequences of arrd-20 and arrd-21 were obtained from Ensembl (http://www.ensembl.org, release 89). The phylogenetic tree of 29 α-arrestins in C. elegans was generated using Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/)20 and re-visualized using the Dendroscope 3 (version 3.5.9)21. For the α-arrestins that have multiple isoforms, isoform a was used for the analysis.
RNAi clones
Twenty one RNAi clones that target C. elegans α-arrestin genes were used from two commercial C. elegans RNAi feeding libraries. Specifically, RNAi clones targeting arrd-2, arrd-6, arrd-7, arrd-8, arrd-9, arrd-10, arrd-13, arrd-16, arrd-18, arrd-23, arrd-24, arrd-25, arrd-28 and ttm-2 (toxin-regulated targets of MAPK 2) were obtained from Ahringer laboratory library (Geneservice Ltd., Cambridge, UK), while those against arrd-1, arrd-3, arrd-4, arrd-5, arrd-14, arrd-15 and arrd-19 were from Vidal laboratory library (Source BioScience, Nottingham, UK). Three RNAi clones targeting arrd-11, arrd-17, and arrd-26 were generated by molecular cloning using infusion recombinase (EZ-FusionTM Cloning Kit, Enzynomics, Daejeon, South Korea). The N2 genomic DNA was obtained through the lysis of worms using proteinase K (Invitrogen, Carlsbad, CA, USA) and N2 complementary DNA was obtained from RNA extraction using RNAiso Plus (Takara, Shiga, Japan) followed by reverse transcription using ImProm-II reverse transcriptase (Promega, Madison, WI, USA, USA). The infusion reaction between PCR products and pL4440 plasmids (Fire lab C. elegans vector kit, 1999) digested with HindIII (New England Biolabs, Ipswich, MA, USA) and Acc65I (New England Biolabs) was followed by transformation of in-house competent E. coli HT115 cells and by selection of positive clones on ampicillin (USB, Santa Clara, CA, USA)-containing LB plates. Primers (CosmoGenetech, Seoul, South Korea) that were used to amplify coding regions of arrd-11 from N2 genomic DNA, and those of arrd-17 and arrd-26 from N2 complementary DNA are as follows: forward primer 5’-GAATTCGATATCAAGCTCCCTCGTGCAAATTAGGAAA-3’ and reverse primer 5’-CTATAGGGCGAATTGGGGTTCCTCCCACTCCATACA-3’ for arrd-11; forward primer 5’-GAATTCGATATCAAGCTATGGTGCAGTTAGATCGTTTTG-3’ and reverse primer 5’-CTATAGGGCGAATTGGTTAATCGGTATAAAATGG-3’ for arrd-17; forward primer 5’-GAATTCGATATCAAGCTATGAAGGTCGATTACTTCG-3’ and reverse primer 5’- CTATAGGGCGAATTGGCTACTTCTCGGAGCCATTTG-3’ for arrd-26. The sequences of all these 24 α-arrestin RNAi clones were confirmed using DNA sequencing (Solgent, Daejeon, South Korea) before lifespan assays. RNAi clone for arrd-12, arrd-22, or arrd-27 was not generated because we were not able to obtain transformed E. coli (HT115) colonies using arrd-12 or arrd-22 infusion reaction products, or the PCR product for arrd-27 genomic or complementary DNA. arrd-20 and arrd-21 are predicted to be pseudogenes (Wormbase, version WS259) and were excluded from our screen. All RNAi clones used in our screen were examined for their potential off-target effects by using Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap)22, and no significant off-target was predicted except arrd-19 RNAi, which may additionally target a predicted pseudogene arrd-21 (Table S1). Experimental validation by qRT-PCR will be necessary to completely exclude the possible off-target effects caused by RNAi clones.
RNAi screen using lifespan assay
The RNAi screen employing lifespan assay was performed as previously described17. Briefly, E. coli HT115 bacteria that expressed specific dsRNAs were cultured overnight at 37°C in LB media containing 50 μg/ml ampicillin (USB). The cultured bacteria were seeded onto nematode growth media plates containing 50 μg/ml ampicillin and incubated overnight at 37°C. The seeded bacteria were treated with 1 mM isopropyl β-D-1-thiogalactopyranoside (Gold Biotechnology, St. Louis, MO, USA) and incubated at room temperature for approximately 24 h to induce dsRNAs. Age-synchronized wild-type N2 and daf-2(e1370) mutant animals were grown on RNAi plates from embryo to L4 stage at 20°C. Worms were then transferred onto RNAi plates containing 5 μM 5-fluoro-2’deoxyuridine (FUdR; Sigma, St. Louis, MO, USA), which prevents eggs from hatching, at young (day 1) adult stage, and transferred again to new FUdR-containing RNAi plates after 1 or 2 days. Eggs laid by ttm-2 RNAi-treated worms hatched more frequently than control eggs on FUdR-containing plates in two independent lifespan experiments. All lifespan assays were performed at 20°C as duplicates by two independent researchers. The survival of worms was determined by gently touching worms with a platinum wire. Worms that did not respond were counted as dead worms and removed from the plates. Worms that crawled off the plates, burrowed into the agar media, or displayed internal hatching or vulval rupture were censored, but included in the statistical analysis. Lifespan data from two independent lifespan experiments within the experimental sets were pooled and used for statistical analysis (see Dataset 1–Dataset 3). OASIS 2 (Online Application for Survival Analysis 2; https://sbi.postech.ac.kr/oasis2)23 was used for the statistical analysis of lifespan results. P values were calculated using long-rank (Mantel-Cox method) test.
Results
Feeding bacteria expressing dsRNA against each of several C. elegans α-arrestins marginally influenced lifespan
We measured the lifespan of wild-type and long-lived daf-2 mutant animals fed with bacteria expressing dsRNA targeting each of 24 of 29 genes encoding putative α-arrestin proteins (Figure 1A). We used daf-16 RNAi that largely suppressed the longevity of daf-2(-) mutants as a positive control (Figure 1B, Figure S1A, Table S2)17,18. We found that feeding bacteria expressing dsRNA against specific α-arrestin genes caused a minor reduction of lifespan in wild-type or in daf-2(-) mutants (Figure 1B, Figure S1B–U, Table S2). Out of the 24 RNAi clones, bacteria expressing dsRNA targeting arrd (arrestin domain protein)-13, arrd-16, arrd-23, arrd-24 or arrd-25 in wild-type, and bacteria expressing dsRNA against arrd-1, arrd-2, arrd-5, arrd-24 or arrd-28 in daf-2(-) mutant animals decreased lifespan by more than 5% (Figure 1B–E, Figures S1B, D, K, Q, R, T, Table S2). Specifically, bacteria expressing dsRNA against arrd-16 decreased lifespan in wild-type by 9%, and bacteria expressing dsRNA against arrd-1 decreased lifespan in daf-2(-) mutants by 7% (Figure 1B and C). In addition, bacteria expressing dsRNA targeting arrd-24 decreased the lifespan of wild-type and daf-2 mutant animals by 11% and 6%, respectively (Figure 1E). In contrast, bacteria expressing dsRNA against arrd-3 increased the lifespan of wild-type animals by 10% (Figure 1F). Overall, feeding C. elegans with bacteria expressing dsRNA against individual α-arrestin genes appears to have minor effects on lifespan.
Figure 1. Feeding bacteria expressing dsRNA targeting each of Caenorhabditis elegans α-arrestins had little effect on lifespan.
(A) A phylogenetic tree showing C. elegans α-arrestin family members. Asterisks (*) indicate α-arrestins that were not examined in this study and number signs (#) indicate predicted pseudogenes. (B) Each circle represents percent mean lifespan change by feeding wild-type (WT) and daf-2(e1370) (daf-2(-)) mutants with bacteria expressing dsRNA against each of α-arrestin genes. Red triangles indicate lifespan changes by daf-16 RNAi bacteria feeding, which was used as a positive control. (C–F) Lifespan curves of WT and daf-2(-) animals fed with dsRNA-expressing bacteria targeting arrd-16 (C), arrd-1 (D), arrd- 24 (E), and arrd-3 (F). See Table S2 for statistical analysis.
Set
Strain
Treatment
Days
At Risk
Dead
Censored
Percent Mortality
S_hat
Var S_hat
SE(S)
Survival Time
Percent Alive
1
N2
Control RNAi
0
240
0
0
0
1
0
0
0
100
1
N2
Control RNAi
1
240
0
0
0
1
0
0
0
100
1
N2
Control RNAi
2
240
0
0
0
1
0
0
0
100
1
N2
Control RNAi
4
240
1
15
0.42
0.996
0
0.004
0.017
99.6
1
N2
Control RNAi
5
224
0
2
0.42
0.996
0
0.004
0.017
99.6
1
N2
Control RNAi
6
222
2
19
1.31
0.987
0
0.008
0.07
98.7
1
N2
Control RNAi
7
201
0
1
1.31
0.987
0
0.008
0.07
98.7
1
N2
Control RNAi
8
200
2
10
2.3
0.977
0
0.01
0.149
97.7
1
N2
Control RNAi
10
188
6
2
5.42
0.946
0
0.016
0.461
94.6
1
N2
Control RNAi
11
180
3
10
7
0.93
0
0.018
0.635
93
1
N2
Control RNAi
12
167
10
1
12.56
0.874
0.001
0.024
1.303
87.4
1
N2
Control RNAi
14
156
44
4
37.23
0.628
0.001
0.036
4.756
62.8
1
N2
Control RNAi
16
108
28
2
53.5
0.465
0.001
0.038
7.36
46.5
1
N2
Control RNAi
18
78
21
1
66.02
0.34
0.001
0.036
9.613
34
1
N2
Control RNAi
20
56
14
1
74.51
0.255
0.001
0.033
11.312
25.5
1
N2
Control RNAi
22
41
5
0
77.62
0.224
0.001
0.032
11.996
22.4
1
N2
Control RNAi
23
36
17
0
88.19
0.118
0.001
0.025
14.426
11.8
1
N2
Control RNAi
24
19
5
0
91.3
0.087
0
0.022
15.172
8.7
1
N2
Control RNAi
25
14
7
0
95.65
0.044
0
0.016
16.26
4.4
1
N2
Control RNAi
26
7
3
0
97.51
0.025
0
0.012
16.745
2.5
1
N2
Control RNAi
28
4
4
0
100
0
0
0
17.441
0
1
N2
daf-16 RNAi
0
240
0
0
0
1
0
0
0
100
1
N2
daf-16 RNAi
1
240
0
0
0
1
0
0
0
100
1
N2
daf-16 RNAi
2
240
0
0
0
1
0
0
0
100
1
N2
daf-16 RNAi
4
240
2
28
0.83
0.992
0
0.006
0.033
99.2
1
N2
daf-16 RNAi
5
210
0
2
0.83
0.992
0
0.006
0.033
99.2
1
N2
daf-16 RNAi
6
208
1
20
1.31
0.987
0
0.008
0.062
98.7
1
N2
daf-16 RNAi
7
187
0
5
1.31
0.987
0
0.008
0.062
98.7
1
N2
daf-16 RNAi
8
182
4
7
3.48
0.965
0
0.013
0.235
96.5
1
N2
daf-16 RNAi
10
171
9
1
8.56
0.914
0
0.021
0.743
91.4
1
N2
daf-16 RNAi
11
161
37
9
29.57
0.704
0.001
0.034
3.055
70.4
1
N2
daf-16 RNAi
12
115
10
2
35.7
0.643
0.001
0.036
3.79
64.3
1
N2
daf-16 RNAi
14
103
48
5
65.66
0.343
0.001
0.037
7.985
34.3
1
N2
daf-16 RNAi
16
50
25
3
82.83
0.172
0.001
0.031
10.732
17.2
1
N2
daf-16 RNAi
18
22
12
0
92.2
0.078
0.001
0.023
12.418
7.8
1
N2
daf-16 RNAi
20
10
7
0
97.66
0.023
0
0.013
13.51
2.3
1
N2
daf-16 RNAi
22
3
3
0
100
0
0
0
14.025
0
1
N2
arrd-1 RNAi
0
240
0
0
0
1
0
0
0
100
1
N2
arrd-1 RNAi
1
240
0
0
0
1
0
0
0
100
1
N2
arrd-1 RNAi
2
240
0
0
0
1
0
0
0
100
1
N2
arrd-1 RNAi
4
240
0
12
0
1
0
0
0
100
1
N2
arrd-1 RNAi
5
228
0
3
0
1
0
0
0
100
1
N2
arrd-1 RNAi
6
225
1
38
0.44
0.996
0
0.004
0.027
99.6
1
N2
arrd-1 RNAi
7
186
0
1
0.44
0.996
0
0.004
0.027
99.6
1
N2
arrd-1 RNAi
8
185
0
12
0.44
0.996
0
0.004
0.027
99.6
1
N2
arrd-1 RNAi
10
173
2
0
1.6
0.984
0
0.009
0.142
98.4
1
N2
arrd-1 RNAi
11
171
17
7
11.38
0.886
0.001
0.024
1.218
88.6
1
N2
arrd-1 RNAi
12
147
5
2
14.39
0.856
0.001
0.027
1.58
85.6
1
N2
arrd-1 RNAi
14
140
36
3
36.41
0.636
0.001
0.037
4.661
63.6
1
N2
arrd-1 RNAi
16
101
25
1
52.15
0.479
0.002
0.039
7.18
47.9
1
N2
arrd-1 RNAi
18
75
26
2
68.74
0.313
0.001
0.037
10.166
31.3
1
N2
arrd-1 RNAi
20
47
10
1
75.39
0.246
0.001
0.034
11.496
24.6
1
N2
arrd-1 RNAi
22
36
10
0
82.22
0.178
0.001
0.031
13.001
17.8
1
N2
arrd-1 RNAi
23
26
14
0
91.8
0.082
0.001
0.022
15.202
8.2
1
N2
arrd-1 RNAi
24
12
6
0
95.9
0.041
0
0.016
16.186
4.1
1
N2
arrd-1 RNAi
25
6
1
0
96.58
0.034
0
0.015
16.357
3.4
1
N2
arrd-1 RNAi
26
5
4
0
99.32
0.007
0
0.007
17.068
0.7
1
N2
arrd-1 RNAi
28
1
1
0
100
0
0
0
17.26
0
1
N2
arrd-2 RNAi
0
240
0
0
0
1
0
0
0
100
1
N2
arrd-2 RNAi
1
240
0
0
0
1
0
0
0
100
1
N2
arrd-2 RNAi
2
240
0
0
0
1
0
0
0
100
1
N2
arrd-2 RNAi
4
240
0
13
0
1
0
0
0
100
1
N2
arrd-2 RNAi
5
227
0
0
0
1
0
0
0
100
1
N2
arrd-2 RNAi
6
227
1
30
0.44
0.996
0
0.004
0.026
99.6
1
N2
arrd-2 RNAi
7
196
0
0
0.44
0.996
0
0.004
0.026
99.6
1
N2
arrd-2 RNAi
8
196
0
18
0.44
0.996
0
0.004
0.026
99.6
1
N2
arrd-2 RNAi
10
178
3
3
2.12
0.979
0
0.011
0.194
97.9
1
N2
arrd-2 RNAi
11
172
16
2
11.22
0.888
0.001
0.024
1.196
88.8
1
N2
arrd-2 RNAi
12
154
4
0
13.53
0.865
0.001
0.026
1.473
86.5
1
N2
arrd-2 RNAi
14
150
36
3
34.28
0.657
0.001
0.036
4.378
65.7
1
N2
arrd-2 RNAi
16
111
22
0
47.31
0.527
0.001
0.038
6.462
52.7
1
N2
arrd-2 RNAi
18
89
28
0
63.88
0.361
0.001
0.037
9.446
36.1
1
N2
arrd-2 RNAi
20
61
20
0
75.73
0.243
0.001
0.033
11.814
24.3
1
N2
arrd-2 RNAi
22
41
7
0
79.87
0.201
0.001
0.031
12.726
20.1
1
N2
arrd-2 RNAi
23
34
21
0
92.3
0.077
0
0.02
15.585
7.7
1
N2
arrd-2 RNAi
24
13
5
0
95.26
0.047
0
0.016
16.296
4.7
1
N2
arrd-2 RNAi
25
8
2
0
96.45
0.036
0
0.014
16.592
3.6
1
N2
arrd-2 RNAi
26
6
4
0
98.82
0.012
0
0.008
17.208
1.2
1
N2
arrd-2 RNAi
28
2
2
0
100
0
0
0
17.539
0
1
N2
arrd-3 RNAi
0
240
0
0
0
1
0
0
0
100
1
N2
arrd-3 RNAi
1
240
0
0
0
1
0
0
0
100
1
N2
arrd-3 RNAi
2
240
0
0
0
1
0
0
0
100
1
N2
arrd-3 RNAi
4
240
0
6
0
1
0
0
0
100
1
N2
arrd-3 RNAi
5
234
0
1
0
1
0
0
0
100
1
N2
arrd-3 RNAi
6
233
2
23
0.86
0.991
0
0.006
0.052
99.1
1
N2
arrd-3 RNAi
7
208
0
0
0.86
0.991
0
0.006
0.052
99.1
1
N2
arrd-3 RNAi
8
208
2
13
1.81
0.982
0
0.009
0.128
98.2
1
N2
arrd-3 RNAi
10
193
2
1
2.83
0.972
0
0.011
0.23
97.2
1
N2
arrd-3 RNAi
11
190
7
0
6.41
0.936
0
0.017
0.623
93.6
1
N2
arrd-3 RNAi
12
183
4
1
8.45
0.915
0
0.02
0.869
91.5
1
N2
arrd-3 RNAi
14
178
27
8
22.34
0.777
0.001
0.03
2.813
77.7
1
N2
arrd-3 RNAi
16
143
20
3
33.2
0.668
0.001
0.034
4.551
66.8
1
N2
arrd-3 RNAi
18
120
25
1
47.12
0.529
0.001
0.037
7.056
52.9
1
N2
arrd-3 RNAi
20
94
18
0
57.24
0.428
0.001
0.037
9.081
42.8
1
N2
arrd-3 RNAi
22
76
18
0
67.37
0.326
0.001
0.035
11.309
32.6
1
N2
arrd-3 RNAi
23
58
32
1
85.37
0.146
0.001
0.026
15.449
14.6
1
N2
arrd-3 RNAi
24
25
10
0
91.22
0.088
0
0.021
16.853
8.8
1
N2
arrd-3 RNAi
25
15
5
0
94.15
0.059
0
0.018
17.585
5.9
1
N2
arrd-3 RNAi
26
10
4
0
96.49
0.035
0
0.014
18.193
3.5
1
N2
arrd-3 RNAi
28
6
5
0
99.41
0.006
0
0.006
19.012
0.6
1
N2
arrd-3 RNAi
30
1
1
0
100
0
0
0
19.188
0
1
N2
arrd-4 RNAi
0
240
0
0
0
1
0
0
0
100
1
N2
arrd-4 RNAi
1
240
0
0
0
1
0
0
0
100
1
N2
arrd-4 RNAi
2
240
0
0
0
1
0
0
0
100
1
N2
arrd-4 RNAi
4
240
0
15
0
1
0
0
0
100
1
N2
arrd-4 RNAi
5
225
0
2
0
1
0
0
0
100
1
N2
arrd-4 RNAi
6
223
1
22
0.45
0.996
0
0.004
0.027
99.6
1
N2
arrd-4 RNAi
7
200
0
0
0.45
0.996
0
0.004
0.027
99.6
1
N2
arrd-4 RNAi
8
200
2
15
1.44
0.986
0
0.008
0.107
98.6
1
N2
arrd-4 RNAi
10
183
4
5
3.6
0.964
0
0.013
0.322
96.4
1
N2
arrd-4 RNAi
11
174
20
3
14.68
0.853
0.001
0.026
1.541
85.3
1
N2
arrd-4 RNAi
12
151
6
2
18.07
0.819
0.001
0.029
1.948
81.9
1
N2
arrd-4 RNAi
14
143
24
4
31.82
0.682
0.001
0.035
3.873
68.2
1
N2
arrd-4 RNAi
16
115
17
3
41.9
0.581
0.001
0.037
5.485
58.1
1
N2
arrd-4 RNAi
18
95
17
1
52.3
0.477
0.001
0.038
7.357
47.7
1
N2
arrd-4 RNAi
20
77
21
0
65.31
0.347
0.001
0.037
9.959
34.7
1
N2
arrd-4 RNAi
22
56
11
0
72.12
0.279
0.001
0.035
11.458
27.9
1
N2
arrd-4 RNAi
23
45
22
0
85.75
0.142
0.001
0.027
14.593
14.2
1
N2
arrd-4 RNAi
24
23
5
0
88.85
0.112
0.001
0.025
15.336
11.2
1
N2
arrd-4 RNAi
25
18
4
0
91.33
0.087
0
0.022
15.956
8.7
1
N2
arrd-4 RNAi
26
14
5
0
94.42
0.056
0
0.018
16.761
5.6
1
N2
arrd-4 RNAi
28
9
8
0
99.38
0.006
0
0.006
18.149
0.6
1
N2
arrd-4 RNAi
30
1
1
0
100
0
0
0
18.335
0
1
N2
arrd-5 RNAi
0
240
0
0
0
1
0
0
0
100
1
N2
arrd-5 RNAi
1
240
0
0
0
1
0
0
0
100
1
N2
arrd-5 RNAi
2
240
0
0
0
1
0
0
0
100
1
N2
arrd-5 RNAi
4
240
0
9
0
1
0
0
0
100
1
N2
arrd-5 RNAi
5
231
0
3
0
1
0
0
0
100
1
N2
arrd-5 RNAi
6
228
5
27
2.19
0.978
0
0.01
0.132
97.8
1
N2
arrd-5 RNAi
7
196
1
1
2.69
0.973
0
0.011
0.167
97.3
1
N2
arrd-5 RNAi
8
194
0
15
2.69
0.973
0
0.011
0.167
97.3
1
N2
arrd-5 RNAi
10
179
3
3
4.32
0.957
0
0.014
0.33
95.7
1
N2
arrd-5 RNAi
11
173
9
7
9.3
0.907
0
0.021
0.877
90.7
1
N2
arrd-5 RNAi
12
157
2
0
10.46
0.895
0
0.022
1.016
89.5
1
N2
arrd-5 RNAi
14
155
41
6
34.14
0.659
0.001
0.036
4.332
65.9
1
N2
arrd-5 RNAi
16
108
19
2
45.73
0.543
0.001
0.038
6.186
54.3
1
N2
arrd-5 RNAi
18
87
25
1
61.32
0.387
0.001
0.038
8.993
38.7
1
N2
arrd-5 RNAi
20
61
30
2
80.34
0.197
0.001
0.031
12.797
19.7
1
N2
arrd-5 RNAi
22
29
9
0
86.44
0.136
0.001
0.027
14.139
13.6
1
N2
arrd-5 RNAi
23
20
11
1
93.9
0.061
0
0.019
15.854
6.1
1
N2
arrd-5 RNAi
24
8
2
0
95.43
0.046
0
0.017
16.22
4.6
1
N2
arrd-5 RNAi
25
6
4
0
98.48
0.015
0
0.011
16.982
1.5
1
N2
arrd-5 RNAi
26
2
1
0
99.24
0.008
0
0.008
17.181
0.8
1
N2
arrd-5 RNAi
28
1
1
0
100
0
0
0
17.394
0
1
daf-2(e1370)
Control RNAi
0
210
0
0
0
1
0
0
0
100
1
daf-2(e1370)
Control RNAi
1
210
0
0
0
1
0
0
0
100
1
daf-2(e1370)
Control RNAi
2
210
0
0
0
1
0
0
0
100
1
daf-2(e1370)
Control RNAi
4
210
0
6
0
1
0
0
0
100
1
daf-2(e1370)
Control RNAi
7
204
0
8
0
1
0
0
0
100
1
daf-2(e1370)
Control RNAi
8
196
0
0
0
1
0
0
0
100
1
daf-2(e1370)
Control RNAi
10
196
0
4
0
1
0
0
0
100
1
daf-2(e1370)
Control RNAi
11
192
0
3
0
1
0
0
0
100
1
daf-2(e1370)
Control RNAi
13
189
0
4
0
1
0
0
0
100
1
daf-2(e1370)
Control RNAi
14
185
1
3
0.54
0.995
0
0.005
0.076
99.5
1
daf-2(e1370)
Control RNAi
16
181
2
1
1.64
0.984
0
0.009
0.252
98.4
1
daf-2(e1370)
Control RNAi
18
178
0
0
1.64
0.984
0
0.009
0.252
98.4
1
daf-2(e1370)
Control RNAi
19
178
1
0
2.19
0.978
0
0.011
0.357
97.8
1
daf-2(e1370)
Control RNAi
20
177
0
6
2.19
0.978
0
0.011
0.357
97.8
1
daf-2(e1370)
Control RNAi
22
171
1
0
2.76
0.972
0
0.012
0.482
97.2
1
daf-2(e1370)
Control RNAi
23
170
0
2
2.76
0.972
0
0.012
0.482
97.2
1
daf-2(e1370)
Control RNAi
25
168
2
17
3.92
0.961
0
0.015
0.772
96.1
1
daf-2(e1370)
Control RNAi
28
149
4
2
6.5
0.935
0
0.019
1.494
93.5
1
daf-2(e1370)
Control RNAi
30
143
0
2
6.5
0.935
0
0.019
1.494
93.5
1
daf-2(e1370)
Control RNAi
31
141
7
0
11.14
0.889
0.001
0.025
2.933
88.9
1
daf-2(e1370)
Control RNAi
33
134
1
0
11.81
0.882
0.001
0.026
3.152
88.2
1
daf-2(e1370)
Control RNAi
34
133
6
1
15.78
0.842
0.001
0.029
4.504
84.2
1
daf-2(e1370)
Control RNAi
36
126
2
1
17.12
0.829
0.001
0.03
4.986
82.9
1
daf-2(e1370)
Control RNAi
37
123
7
1
21.84
0.782
0.001
0.033
6.731
78.2
1
daf-2(e1370)
Control RNAi
38
115
4
1
24.56
0.754
0.001
0.035
7.764
75.4
1
daf-2(e1370)
Control RNAi
40
110
10
0
31.42
0.686
0.001
0.038
10.507
68.6
1
daf-2(e1370)
Control RNAi
41
100
3
0
33.47
0.665
0.001
0.038
11.351
66.5
1
daf-2(e1370)
Control RNAi
43
97
12
0
41.7
0.583
0.002
0.04
14.89
58.3
1
daf-2(e1370)
Control RNAi
44
85
6
1
45.82
0.542
0.002
0.041
16.701
54.2
1
daf-2(e1370)
Control RNAi
46
78
23
0
61.79
0.382
0.002
0.04
24.05
38.2
1
daf-2(e1370)
Control RNAi
49
55
26
0
79.86
0.201
0.001
0.033
32.9
20.1
1
daf-2(e1370)
Control RNAi
51
29
10
0
86.8
0.132
0.001
0.028
36.442
13.2
1
daf-2(e1370)
Control RNAi
52
19
6
0
90.97
0.09
0.001
0.024
38.61
9
1
daf-2(e1370)
Control RNAi
54
13
5
0
94.44
0.056
0
0.019
40.485
5.6
1
daf-2(e1370)
Control RNAi
55
8
3
0
96.53
0.035
0
0.015
41.631
3.5
1
daf-2(e1370)
Control RNAi
57
5
2
0
97.92
0.021
0
0.012
42.423
2.1
1
daf-2(e1370)
Control RNAi
60
3
2
0
99.31
0.007
0
0.007
43.257
0.7
1
daf-2(e1370)
Control RNAi
63
1
1
0
100
0
0
0
43.694
0
1
daf-2(e1370)
daf-16 RNAi
0
240
0
0
0
1
0
0
0
100
1
daf-2(e1370)
daf-16 RNAi
1
240
0
0
0
1
0
0
0
100
1
daf-2(e1370)
daf-16 RNAi
2
240
0
0
0
1
0
0
0
100
1
daf-2(e1370)
daf-16 RNAi
4
240
0
3
0
1
0
0
0
100
1
daf-2(e1370)
daf-16 RNAi
7
237
4
12
1.69
0.983
0
0.008
0.118
98.3
1
daf-2(e1370)
daf-16 RNAi
8
221
0
1
1.69
0.983
0
0.008
0.118
98.3
1
daf-2(e1370)
daf-16 RNAi
10
220
4
2
3.48
0.965
0
0.012
0.297
96.5
1
daf-2(e1370)
daf-16 RNAi
11
214
4
1
5.28
0.947
0
0.015
0.495
94.7
1
daf-2(e1370)
daf-16 RNAi
13
209
2
2
6.19
0.938
0
0.016
0.613
93.8
1
daf-2(e1370)
daf-16 RNAi
14
205
8
2
9.85
0.902
0
0.02
1.126
90.2
1
daf-2(e1370)
daf-16 RNAi
16
195
11
1
14.93
0.851
0.001
0.024
1.939
85.1
1
daf-2(e1370)
daf-16 RNAi
18
183
10
0
19.58
0.804
0.001
0.027
2.776
80.4
1
daf-2(e1370)
daf-16 RNAi
19
173
16
0
27.02
0.73
0.001
0.03
4.189
73
1
daf-2(e1370)
daf-16 RNAi
20
157
18
3
35.39
0.646
0.001
0.032
5.863
64.6
1
daf-2(e1370)
daf-16 RNAi
22
136
17
1
43.46
0.565
0.001
0.034
7.64
56.5
1
daf-2(e1370)
daf-16 RNAi
23
118
33
0
59.27
0.407
0.001
0.034
11.276
40.7
1
daf-2(e1370)
daf-16 RNAi
25
85
30
0
73.65
0.264
0.001
0.03
14.87
26.4
This is a portion of the data; to view all the data, please download the file.
Dataset 1.Kaplan-Meier estimator of RNAi lifespan experiments.
Kaplan-Meier estimate values were calculated from pooled lifespan data of two independent lifespan experiments using OASIS2 (https://sbi.postech.ac.kr/oasis2/). ‘At risk’ indicates the number of individuals at risk just prior to a specific time point. ‘S_hat’ indicates Kaplan-Meier estimate of survival function, ‘Var S_hat’ indicates variance of ‘S_hat’, and ‘SE(S)’ indicates standard error of ‘S_hat’.
Set
Strain
Treatment
Number of subjects
Mean survival days
Standard error of mean survival
95 percent confidence interval of mean
Age in days at 25 percent mortailty
Age in days at 50 percent mortailty
Age in days at 75 percent mortailty
Age in days at 90 percent mortailty
Age in days at 100 percent mortailty
95 percent median confidence interval
25 percent linear interpolation of mortality curve
50 percent linear interpolation of mortality curve
75 percent linear interpolation of mortality curve
90 percent linear interpolation of mortality curve
100 percent linear interpolation of mortality curve
1
N2
control RNAi
240
17.44
0.35
16.75 ~ 18.13
14
16
22
24
28
16.0 ~ 16.0
13.01
15.57
20.31
23.58
28
1
N2
daf-16 RNAi
240
14.03
0.25
13.54 ~ 14.51
11
14
16
18
22
- ~ -
10.78
12.95
15.09
17.53
22
1
N2
arrd-1 RNAi
240
17.26
0.34
16.60 ~ 17.92
14
16
20
23
28
16.0 ~ 16.0
12.96
15.73
19.88
22.81
28
1
N2
arrd-2 RNAi
240
17.54
0.33
16.89 ~ 18.19
14
18
20
23
28
16.0 ~ 16.0
13.11
16.32
19.88
22.81
28
1
N2
arrd-3 RNAi
240
19.19
0.34
18.53 ~ 19.85
16
20
23
24
30
18.0 ~ 18.0
14.49
18.57
22.42
23.79
30
1
N2
arrd-4 RNAi
240
18.34
0.39
17.56 ~ 19.11
14
18
23
25
30
18.0 ~ 18.0
13.01
17.56
22.21
24.46
30
1
N2
arrd-5 RNAi
240
17.39
0.32
16.76 ~ 18.02
14
18
20
23
28
16.0 ~ 16.0
13.23
16.55
19.44
22.48
28
1
daf-2(e1370)
control RNAi
210
43.69
0.69
42.35 ~ 45.04
40
46
49
52
63
44.0 ~ 44.0
38.13
44.52
48.19
51.77
63
1
daf-2(e1370)
daf-16 RNAi
240
22.52
0.37
21.79 ~ 23.25
19
23
28
28
34
22.0 ~ 22.0
18.73
22.41
25.22
27.63
34
1
daf-2(e1370)
arrd-1 RNAi
180
40.81
0.85
39.15 ~ 42.47
36
43
46
52
60
41.0 ~ 44.0
34.76
42.49
45.88
51.34
60
1
daf-2(e1370)
arrd-2 RNAi
210
41.47
0.66
40.18 ~ 42.76
36
44
46
51
57
43.0 ~ 43.0
35.75
43.17
45.9
50.15
57
1
daf-2(e1370)
arrd-3 RNAi
240
43.32
0.65
42.04 ~ 44.59
38
46
49
52
63
43.0 ~ 44.0
37.42
44.03
48.52
51.77
63
1
daf-2(e1370)
arrd-4 RNAi
240
41.74
0.65
40.45 ~ 43.02
37
43
49
52
60
41.0 ~ 43.0
36.34
42.36
47.09
51.55
60
1
daf-2(e1370)
arrd-5 RNAi
240
41.11
0.6
39.93 ~ 42.28
37
43
46
51
57
41.0 ~ 43.0
36.08
41.49
45.81
50.26
57
2
N2
control RNAi
240
21.66
0.35
20.97 ~ 22.36
18
22
24
27
34
21.0 ~ 21.0
17.01
21
23.94
26.51
34
2
N2
daf-16 RNAi
240
17.63
0.31
17.02 ~ 18.24
15
18
22
24
27
18.0 ~ 18.0
12.51
16.89
21.09
22.51
27
2
N2
arrd-6 RNAi
240
20.51
0.43
19.68 ~ 21.35
15
19
24
30
37
19.0 ~ 21.0
14.27
18.98
23.93
27.15
37
2
N2
arrd-7 RNAi
240
20.92
0.39
20.17 ~ 21.68
15
21
24
27
34
19.0 ~ 21.0
14.77
19.8
23.81
26.85
34
2
N2
arrd-8 RNAi
240
20.52
0.42
19.69 ~ 21.35
15
21
24
30
30
19.0 ~ 21.0
13.94
19.31
23.77
27.72
30
2
N2
arrd-9 RNAi
240
20.77
0.39
20.00 ~ 21.54
15
21
24
27
34
19.0 ~ 19.0
14.71
19.27
23.85
26.91
34
2
N2
arrd-10 RNAi
240
20.22
0.41
19.42 ~ 21.02
15
21
24
27
30
19.0 ~ 21.0
13.79
19.6
23.46
26.53
30
2
daf-2(e1370)
control RNAi
240
44.31
0.73
42.88 ~ 45.75
39
48
51
57
62
44.0 ~ 45.0
37.78
45.29
50.37
55.18
62
2
daf-2(e1370)
daf-16 RNAi
241
25.28
0.41
24.47 ~ 26.09
21
24
30
34
40
24.0 ~ 24.0
20.6
23.58
27.85
33.09
40
2
daf-2(e1370)
arrd-6 RNAi
240
44.25
0.66
42.95 ~ 45.54
39
45
51
55
62
44.0 ~ 45.0
38.29
44.79
49.25
54.51
62
2
daf-2(e1370)
arrd-7 RNAi
240
43.09
0.74
41.64 ~ 44.55
37
45
51
55
62
44.0 ~ 45.0
36.15
44.52
49.54
54.84
62
2
daf-2(e1370)
arrd-8 RNAi
240
43.33
0.66
42.04 ~ 44.61
39
45
48
54
62
44.0 ~ 45.0
38.9
44.06
47.99
52.18
62
2
daf-2(e1370)
arrd-9 RNAi
240
43.59
0.66
42.30 ~ 44.88
40
45
51
54
62
44.0 ~ 45.0
39.41
44.68
48.43
52.92
62
2
daf-2(e1370)
arrd-10 RNAi
240
46.02
0.75
44.56 ~ 47.48
40
48
55
59
66
45.0 ~ 45.0
39.45
46.23
54.18
57.44
66
3
N2
control RNAi
240
19.83
0.32
19.21 ~ 20.45
17
20
24
24
34
- ~ -
16.35
19.25
22.42
23.98
34
3
N2
daf-16 RNAi
240
14.82
0.22
14.40 ~ 15.25
12
14
17
18
22
14.0 ~ 14.0
11.53
13.92
16.71
17.98
22
3
N2
arrd-13 RNAi
210
18.67
0.3
18.08 ~ 19.27
16
18
22
24
30
18.0 ~ 18.0
15.38
17.96
20.24
23.29
30
3
N2
arrd-14 RNAi
240
19.13
0.3
18.53 ~ 19.72
16
20
22
24
28
18.0 ~ 18.0
15.93
18.58
21.53
23.73
28
3
N2
arrd-15 RNAi
240
18.86
0.32
18.23 ~ 19.49
14
20
22
26
30
18.0 ~ 18.0
13.76
18.24
20.83
24.95
30
3
N2
arrd-16 RNAi
240
17.98
0.31
17.38 ~ 18.58
14
17
20
24
28
17.0 ~ 17.0
13.36
16.89
19.81
23.92
28
3
N2
arrd-18 RNAi
240
18.89
0.33
18.25 ~ 19.54
14
18
22
26
32
17.0 ~ 18.0
13.89
17.94
21.14
25.05
32
3
daf-2(e1370)
control RNAi
210
44.89
0.85
43.23 ~ 46.55
38
47
53
58
71
44.0 ~ 47.0
37.86
45.39
52.85
57.13
71
3
daf-2(e1370)
daf-16 RNAi
240
23.45
0.33
22.80 ~ 24.11
20
24
26
28
38
24.0 ~ 24.0
18.94
23.2
25.67
27.83
38
3
daf-2(e1370)
arrd-13 RNAi
210
45.69
0.9
43.92 ~ 47.46
40
47
53
59
70
44.0 ~ 47.0
38.05
46.07
52.35
57.43
70
3
daf-2(e1370)
arrd-14 RNAi
240
42.94
0.84
41.30 ~ 44.58
37
44
50
56
67
44.0 ~ 44.0
36.09
43.51
49.94
54.3
67
3
daf-2(e1370)
arrd-15 RNAi
240
45.79
0.67
44.47 ~ 47.11
41
47
51
56
67
- ~ -
40.15
45.44
50.73
55.53
67
3
daf-2(e1370)
arrd-16 RNAi
240
43.29
0.82
41.68 ~ 44.89
37
47
51
56
64
44.0 ~ 44.0
36.65
44.82
50.49
54.23
64
3
daf-2(e1370)
arrd-18 RNAi
180
45.1
0.95
43.24 ~ 46.96
38
47
53
59
64
44.0 ~ 47.0
37.63
45.44
52.26
58.54
64
4
N2
control RNAi
240
18.04
0.28
17.49 ~ 18.59
15
18
21
23
29
17.0 ~ 18.0
14.13
17.43
20.28
22.45
29
4
N2
daf-16 RNAi
240
14.85
0.21
14.44 ~ 15.26
12
15
17
18
23
14.0 ~ 14.0
11.73
14.24
16.38
17.98
23
4
N2
arrd-23 RNAi
240
16.78
0.3
16.19 ~ 17.38
14
17
19
23
27
15.0 ~ 17.0
13.03
16.15
18.99
21.81
27
4
N2
arrd-24 RNAi
240
16.05
0.22
15.61 ~ 16.49
14
15
18
21
26
15.0 ~ 15.0
12.82
14.91
17.16
19.85
26
4
N2
arrd-25 RNAi
240
16.86
0.27
16.32 ~ 17.39
15
17
19
21
26
- ~ -
12.94
15.94
18.56
20.74
26
4
N2
arrd-26 RNAi
240
17.22
0.28
16.66 ~ 17.78
15
17
19
23
29
15.0 ~ 15.0
12.86
15.37
18.84
22.7
29
4
N2
arrd-28 RNAi
240
17.53
0.27
17.00 ~ 18.07
15
17
21
23
29
17.0 ~ 17.0
13.4
16.79
19.31
21.92
29
4
daf-2(e1370)
control RNAi
210
45.93
0.75
44.45 ~ 47.40
39
50
53
56
61
49.0 ~ 49.0
38.88
49.03
52.68
55.15
61
4
daf-2(e1370)
daf-16 RNAi
240
20.29
0.35
19.60 ~ 20.97
18
21
24
27
29
18.0 ~ 18.0
15.32
18.75
23.43
26.7
29
4
daf-2(e1370)
arrd-23 RNAi
240
44.15
0.7
42.78 ~ 45.51
37
47
52
56
60
46.0 ~ 47.0
36.8
46.71
51.15
53.44
60
4
daf-2(e1370)
arrd-24 RNAi
211
43
0.75
41.52 ~ 44.47
39
46
50
53
59
46.0 ~ 46.0
38.29
44.82
49.36
52.81
59
4
daf-2(e1370)
arrd-25 RNAi
240
43.74
0.64
42.47 ~ 45.00
40
46
50
53
59
46.0 ~ 46.0
39.53
45.97
49.47
52.41
59
4
daf-2(e1370)
arrd-26 RNAi
241
44.67
0.64
43.42 ~ 45.93
39
47
52
53
59
46.0 ~ 47.0
38.39
46.61
50.32
52.94
59
4
daf-2(e1370)
arrd-28 RNAi
150
43.52
0.85
41.85 ~ 45.19
37
47
50
53
56
46.0 ~ 47.0
36.86
46.13
49.65
52.53
56
5
N2
control RNAi
240
20.31
0.43
19.47 ~ 21.16
15
21
24
28
38
19.0 ~ 19.0
15
19.76
23.86
27.2
38
5
N2
daf-16 RNAi
240
15.63
0.35
14.95 ~ 16.31
12
15
18
22
27
14.0 ~ 15.0
10.87
14.45
17.88
21.66
27
5
N2
arrd-11 RNAi
240
20.17
0.42
19.35 ~ 20.99
17
21
25
27
40
19.0 ~ 19.0
15.29
19.65
24.05
26.39
40
5
N2
arrd-17 RNAi
240
20.52
0.41
19.72 ~ 21.33
17
21
24
27
40
19.0 ~ 19.0
16.22
19.65
23.16
26.51
40
5
N2
arrd-19 RNAi
240
19.56
0.41
18.75 ~ 20.36
15
19
24
27
36
18.0 ~ 19.0
14.6
18.7
23.23
25.9
36
5
N2
ttm-2 RNAi
240
19.11
0.44
18.24 ~ 19.97
15
19
24
27
36
18.0 ~ 19.0
14.28
18.07
22.39
26.62
36
5
daf-2(e1370)
control RNAi
240
42.97
0.7
41.60 ~ 44.33
38
46
50
54
61
42.0 ~ 42.0
36.55
42.74
49.38
53.12
61
5
daf-2(e1370)
daf-16 RNAi
243
21.54
0.37
20.82 ~ 22.27
17
21
25
28
38
21.0 ~ 21.0
16.88
20.73
24.2
27.62
38
5
daf-2(e1370)
arrd-11 RNAi
210
42.18
0.72
40.77 ~ 43.59
38
42
50
54
58
42.0 ~ 42.0
35.87
41.78
49.22
52.53
58
5
daf-2(e1370)
arrd-17 RNAi
240
41.32
0.62
40.09 ~ 42.54
38
42
46
54
59
- ~ -
35.57
40.07
45.46
52.16
59
5
daf-2(e1370)
arrd-19 RNAi
240
41.1
0.59
39.94 ~ 42.26
38
42
46
50
59
- ~ -
35.72
40.72
44.68
49.62
59
Dataset 2.Mean lifespan and mortality rates of RNAi experiment results.
Mean lifespan, ages at different percent mortality, linear interpolation of mortality curve at specific mortality rate were calculated from pooled lifespan data of two independent lifespan experiments. Data were obtained using OASIS2 (https://sbi.postech.ac.kr/oasis2/).
Set
Condition 1 strain
Condition 1 treatment
Condition 2 strain
Condition 2 treatment
Chi square
P value
Bonferroni P value
1
N2
control RNAi
N2
daf-16 RNAi
60.04
0
0
1
N2
control RNAi
N2
arrd-1 RNAi
0.69
0.4068
1
1
N2
control RNAi
N2
arrd-2 RNAi
0.08
0.7799
1
1
N2
control RNAi
N2
arrd-3 RNAi
8.09
0.0045
0.0268
1
N2
control RNAi
N2
arrd-4 RNAi
3.07
0.0798
0.4786
1
N2
control RNAi
N2
arrd-5 RNAi
0.47
0.4941
1
1
daf-2(e1370)
control RNAi
daf-2(e1370)
daf-16 RNAi
342.23
0
0
1
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-1 RNAi
4.87
0.0274
0.1642
1
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-2 RNAi
6.39
0.0115
0.0344
1
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-3 RNAi
0
0.9633
1
1
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-4 RNAi
2.54
0.1107
0.6644
1
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-5 RNAi
9.05
0.0026
0.0157
2
N2
control RNAi
N2
daf-16 RNAi
60.55
0
0
2
N2
control RNAi
N2
arrd-6 RNAi
1.41
0.2356
1
2
N2
control RNAi
N2
arrd-7 RNAi
0.71
0.3993
1
2
N2
control RNAi
N2
arrd-8 RNAi
0.94
0.3324
1
2
N2
control RNAi
N2
arrd-9 RNAi
1.23
0.2675
1
2
N2
control RNAi
N2
arrd-10 RNAi
3.24
0.072
0.432
2
daf-2(e1370)
control RNAi
daf-2(e1370)
daf-16 RNAi
311.97
0
0
2
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-6 RNAi
0.57
0.4514
1
2
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-7 RNAi
0.59
0.443
1
2
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-8 RNAi
3.79
0.0516
0.3099
2
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-9 RNAi
3.22
0.0729
0.4376
2
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-10 RNAi
5.82
0.0158
0.0948
3
N2
control RNAi
N2
daf-16 RNAi
158.43
0
0
3
N2
control RNAi
N2
arrd-13
8.81
0.003
0.018
3
N2
control RNAi
N2
arrd-14
3.81
0.0509
0.3053
3
N2
control RNAi
N2
arrd-15
3
0.0831
0.4983
3
N2
control RNAi
N2
arrd-16
15.5
0.0001
0.0005
3
N2
control RNAi
N2
arrd-18
2.2
0.1381
0.8287
3
daf-2(e1370)
control RNAi
daf-2(e1370)
daf-16 RNAi
309.08
0
0
3
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-13
0.54
0.4607
1
3
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-14
4.09
0.0433
0.2596
3
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-15
0.02
0.8875
1
3
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-16
2.45
0.1174
0.7042
3
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-18
0.49
0.486
1
4
N2
control RNAi
N2
daf-16 RNAi
85.14
0
0
4
N2
control RNAi
N2
arrd-23 RNAi
7.03
0.008
0.0481
4
N2
control RNAi
N2
arrd-24 RNAi
33.81
0.000000006
0.000000036
4
N2
control RNAi
N2
arrd-25 RNAi
9.4
0.0022
0.013
4
N2
control RNAi
N2
arrd-26 RNAi
2.24
0.1342
0.8053
4
N2
control RNAi
N2
arrd-28 RNAi
1.51
0.2188
1
4
daf-2(e1370)
control RNAi
daf-2(e1370)
daf-16 RNAi
392.76
0
0
4
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-23 RNAi
8.34
0.0039
0.0233
4
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-24 RNAi
20.6
0.0000056
0.000034
4
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-25 RNAi
24.9
0.00000061
0.0000036
4
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-26 RNAi
11.75
0.0006
0.0037
4
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-28 RNAi
17.71
0.000026
0.0002
5
N2
control RNAi
N2
daf-16 RNAi
67.47
0
0
5
N2
control RNAi
N2
arrd-11 RNAi
0.08
0.7707
1
5
N2
control RNAi
N2
arrd-17 RNAi
0
0.9914
1
5
N2
control RNAi
N2
arrd-19 RNAi
1.72
0.1891
0.9455
5
N2
control RNAi
N2
ttm-2 RNAi
3.22
0.0729
0.3644
5
daf-2(e1370)
control RNAi
daf-2(e1370)
daf-16 RNAi
368.1
0
0
5
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-11 RNAi
1.01
0.3156
1
5
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-17 RNAi
5.85
0.0156
0.078
5
daf-2(e1370)
control RNAi
daf-2(e1370)
arrd-19 RNAi
11.49
0.0007
0.0035
Dataset 3.Statistical analysis of lifespan data.
Statistical test results (Chi square, p value, Bonferroni p value) were calculated between ‘condition 1’ and ‘condition 2’. Test results were obtained using OASIS2 (https://sbi.postech.ac.kr/oasis2/).
Discussion
The lifespan-regulatory functions of α-arrestins remain largely unexplored at the organism level. Here we showed that feeding bacteria expressing dsRNA targeting individual C. elegans α-arrestins had little effect on lifespan in wild-type or daf-2 mutants. Our study has limitations that need to be considered for interpretation. First, feeding RNAi targeting some α-arrestins might be insufficient for causing strong lifespan phenotypes. This may be because many of C. elegans α-arrestins are predicted to be expressed in neurons7,9,24–26, which are refractory to RNAi27–29. Lifespan assays using RNAi-hypersensitive mutants, including rrf-3(-) and eri-1(-) animals29–31, treated with α-arrestin RNAi, or using α-arrestin null mutants will help address this issue. In addition, as we did not test whether RNAi targeting each α-arrestin gene was effective by using quantitative RT-PCR, our negative data should be interpreted with caution. Second, C. elegans α-arrestins may have functional redundancy, considering the large number of the α-arrestin family members in C. elegans and their sequence similarity1,7, which may obscure examining the functional importance of each α-arrestin. In addition, some α-arrestins may mostly function by modulating the action of their interacting proteins6. In this case, genetic inhibition of α-arrestins may rather subtly affect the functions of their interacting partners that directly regulate physiology, such as aging and longevity, causing weak or no phenotypes. Thus, it will be interesting to test the effects of simultaneous inhibition of α-arrestins possibly through targeting the arrestin domain, and to identify and to functionally characterize proteins that bind C. elegans α-arrestins. Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.
In mammals, several α-arrestins are implicated in metabolic regulation5. TXNIP (thioredoxin-interacting protein), an inhibitor of thioredoxin in mammals32–34, is a crucial negative regulator of glucose uptake35,36. ARRDC4 inhibits glucose uptake in cultured mammalian cells as well36, and ARRDC3 deficiency protects against obesity in male mice through increasing energy expenditure37. Because metabolism is closely associated with aging38, it will be interesting to test whether α-arrestins in complex metazoans play roles in aging via regulating metabolism.
Data availability
Dataset 1. Kaplan-Meier estimator of RNAi lifespan experiments. Kaplan-Meier estimate values were calculated from pooled lifespan data of two independent lifespan experiments using OASIS2 (https://sbi.postech.ac.kr/oasis2/). ‘At risk’ indicates the number of individuals at risk just prior to a specific time point. ‘S_hat’ indicates Kaplan-Meier estimate of survival function, ‘Var S_hat’ indicates variance of ‘S_hat’, and ‘SE(S)’ indicates standard error of ‘S_hat’. doi, 10.5256/f1000research.12337.d17315839
Dataset 2. Mean lifespan and mortality rates of RNAi experiment results. Mean lifespan, ages at different percent mortality, linear interpolation of mortality curve at specific mortality rate were calculated from pooled lifespan data of two independent lifespan experiments. Data were obtained using OASIS2 (https://sbi.postech.ac.kr/oasis2/). doi, 10.5256/f1000research.12337.d17315940
Dataset 3. Statistical analysis of lifespan data. Statistical test results (Chi square, p value, Bonferroni p value) were calculated between ‘condition 1’ and ‘condition 2’. Test results were obtained using OASIS2 (https://sbi.postech.ac.kr/oasis2/). doi, 10.5256/f1000research.12337.d17316041
Competing interests
No competing interests were disclosed.
Grant information
This work was supported by the Korean Government (MSIP) through the National Research Foundation of Korea (NRF) [NRF-2016R1E1A1A01941152] funded to S-J.V.L.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgments
We thank the Lee laboratory members and Dr. Young Kwon for critical comments on the manuscript.
Supplementary material
Figure S1. Feeding bacteria expressing dsRNA against each Caenorhabditis elegans α-arrestin had minor effects on the lifespan of wild-type or daf-2(-) mutants. (A–U). Lifespan curves of wild-type (WT) and daf-2(e1370) (daf-2(-)) mutants fed with bacteria expressing dsRNA targeting daf-16 (A), which was used as a positive control, arrd-2 (B), arrd-4 (C), arrd-5 (D), arrd-6 (E), arrd-7 (F), arrd-8 (G), arrd-9 (H), arrd-10 (I), arrd-11 (J), arrd-13 (K), arrd-14 (L), arrd-15 (M), arrd-17 (N), arrd-18 (O), arrd-19 (P), arrd-23 (Q), arrd-25 (R), arrd-26 (S), arrd-28 (T) and ttm-2 (U). See Table S2 for statistical analysis and for additional repeats of lifespan data using daf-16 RNAi.
Table S1. Possible RNAi target prediction. RNAi clones used in the screen were examined for their possible off-targets by Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap)22. For RNAi clone names, ‘mv’ indicates an RNAi clone from Vidal library and ‘sjj’ indicates an RNAi clone from Ahringer library. RNAi clones generated by us for this study are labeled as ‘Lee lab’. Scores for potential target transcripts were calculated as described previously22 and were rounded off to the second digit after the decimal point.
Table S2. Lifespan data of Caenorhabditis elegans fed with α-arrestin dsRNA-expressing bacteria. Lifespan data within the solid lines indicate the pooled experimental datasets that are performed at the same time by two independent researchers. P values were calculated within the sets using log-rank (Mantel-Cox) method23. Percent (%) changes and p values for the lifespan data of wild-type (WT) and daf-2(e1370) worms fed with specific α-arrestin dsRNA-expressing bacteria were calculated against those of WT and daf-2(e1370) worms fed with control dsRNA-expressing bacteria within the sets, respectively. Percent (%) changes and p values for the lifespan data of daf-2(e1370) mutants fed with control dsRNA-expressing bacteria were calculated against those of WT worms fed with control dsRNA-expressing bacteria. SEM represents standard error of the mean, and 75th percentile indicates the age (days) at 75% mortality.
3.
Polekhina G, Ascher DB, Kok SF, et al.:
Structure of the N-terminal domain of human thioredoxin-interacting protein.
Acta Crystallogr D Biol Crystallogr.
2013; 69(Pt 3): 333–344. PubMed Abstract
| Publisher Full Text
4.
Becuwe, M, Herrador A, Haguenauer-Tsapis R, et al.:
Ubiquitin-mediated regulation of endocytosis by proteins of the arrestin family.
Biochem Res Int.
2012; 2012: 242764. PubMed Abstract
| Publisher Full Text
| Free Full Text
6.
Puca L, Brou C:
Α-arrestins - new players in Notch and GPCR signaling pathways in mammals.
J Cell Sci.
2014; 127(Pt 7): 1359–1367. PubMed Abstract
| Publisher Full Text
8.
Huffman DL, Abrami L, Sasik R, et al.:
Mitogen-activated protein kinase pathways defend against bacterial pore-forming toxins.
Proc Natl Acad Sci U S A.
2004; 101(30): 10995–11000. PubMed Abstract
| Publisher Full Text
| Free Full Text
9.
Jee C, Choi TW, Kalichamy K, et al.:
CNP-1 (ARRD-17), a novel substrate of calcineurin, is critical for modulation of egg-laying and locomotion in response to food and lysine sensation in Caenorhabditis elegans.
J Mol Biol.
2012; 417(3): 165–178. PubMed Abstract
| Publisher Full Text
11.
An SWA, Artan M, Park S, et al.:
Longevity Regulation by Insulin/IGF-1 Signalling. In Ageing: Lessons from C. elegans. A. Olsen and M.S. Gill, Editors. Springer International Publishing, 2017; 63–81. Publisher Full Text
12.
Altintas O, Park S, Lee SJ:
The role of insulin/IGF-1 signaling in the longevity of model invertebrates, C. elegans and D. melanogaster.
BMB Rep.
2016; 49(2): 81–92. PubMed Abstract
| Publisher Full Text
| Free Full Text
14.
Lee SJ, Hwang AB, Kenyon C:
Inhibition of respiration extends C. elegans life span via reactive oxygen species that increase HIF-1 activity.
Curr Biol.
2010; 20(23): 2131–2136. PubMed Abstract
| Publisher Full Text
| Free Full Text
15.
Hwang W, Artan M, Seo M, et al.:
Inhibition of elongin C promotes longevity and protein homeostasis via HIF-1 in C. elegans.
Aging Cell.
2015; 14(6): 995–1002. PubMed Abstract
| Publisher Full Text
| Free Full Text
16.
Lee D, Jeong DE, Son HG, et al.:
SREBP and MDT-15 protect C. elegans from glucose-induced accelerated aging by preventing accumulation of saturated fat.
Genes Dev.
2015; 29(23): 2490–2503. PubMed Abstract
| Publisher Full Text
| Free Full Text
17.
Seo M, Seo K, Hwang W, et al.:
RNA helicase HEL-1 promotes longevity by specifically activating DAF-16/FOXO transcription factor signaling in Caenorhabditis elegans.
Proc Natl Acad Sci U S A.
2015; 112(31): E4246–E4255. PubMed Abstract
| Publisher Full Text
| Free Full Text
18.
Samuelson AV, Carr CE, Ruvkun G:
Gene activities that mediate increased life span of C. elegans insulin-like signaling mutants.
Genes Dev.
2007; 21(22): 2976–2994. PubMed Abstract
| Publisher Full Text
| Free Full Text
21.
Huson DH, Scornavacca C:
Dendroscope 3: an interactive tool for rooted phylogenetic trees and networks.
Syst Biol.
2012; 61(6): 1061–1067. PubMed Abstract
| Publisher Full Text
22.
Thakur N, Pujol N, Tichit L, et al.:
Clone mapper: an online suite of tools for RNAi experiments in Caenorhabditis elegans.
G3 (Bethesda).
2014; 4(11): 2137–2145. PubMed Abstract
| Publisher Full Text
| Free Full Text
23.
Han SK, Lee D, Lee H, et al.:
OASIS 2: online application for survival analysis 2 with features for the analysis of maximal lifespan and healthspan in aging research.
Oncotarget.
2016; 7(35): 56147–56152. PubMed Abstract
| Publisher Full Text
| Free Full Text
24.
Kunitomo H, Uesugi H, Kohara Y, et al.:
Identification of ciliated sensory neuron-expressed genes in Caenorhabditis elegans using targeted pull-down of poly(A) tails.
Genome Biol.
2005; 6(2): R17. PubMed Abstract
| Publisher Full Text
| Free Full Text
25.
Lynch AS, Briggs D, Hope IA:
Developmental expression pattern screen for genes predicted in the C. elegans genome sequencing project.
Nat Genet.
1995; 11(3): 309–313. PubMed Abstract
| Publisher Full Text
26.
Mounsey A, Bauer P, Hope IA:
Evidence suggesting that a fifth of annotated Caenorhabditis elegans genes may be pseudogenes.
Genome Res.
2002; 12(5): 770–775. PubMed Abstract
| Free Full Text
27.
Timmons L, Court DL, Fire A:
Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans.
Gene.
2001; 263(1–2): 103–112. PubMed Abstract
| Publisher Full Text
28.
Kamath RS, Martinez-Campos M, Zipperlen P, et al.:
Effectiveness of specific RNA-mediated interference through ingested double-stranded RNA in Caenorhabditis elegans.
Genome Biol.
2001; 2(1): RESEARCH0002. PubMed Abstract
| Publisher Full Text
| Free Full Text
29.
Kennedy S, Wang D, Ruvkun G:
A conserved siRNA-degrading RNase negatively regulates RNA interference in C. elegans.
Nature.
2004; 427(6975): 645–649. PubMed Abstract
| Publisher Full Text
30.
Simmer F, Tijsterman M, Parrish S, et al.:
Loss of the putative RNA-directed RNA polymerase RRF-3 makes C. elegans hypersensitive to RNAi.
Curr Biol.
2002; 12(15): 1317–1319. PubMed Abstract
| Publisher Full Text
31.
Sijen T, Fleenor J, Simmer F, et al.:
On the role of RNA amplification in dsRNA-triggered gene silencing.
Cell.
2001; 107(4): 465–476. PubMed Abstract
| Publisher Full Text
32.
Junn E, Han SH, Im JY, et al.:
Vitamin D3 up-regulated protein 1 mediates oxidative stress via suppressing the thioredoxin function.
J Immunol.
2000; 164(12): 6287–6295. PubMed Abstract
| Publisher Full Text
33.
Nishiyama A, Matsui M, Iwata S, et al.:
Identification of thioredoxin-binding protein-2/vitamin D3 up-regulated protein 1 as a negative regulator of thioredoxin function and expression.
J Biol Chem.
1999; 274(31): 21645–21650. PubMed Abstract
| Publisher Full Text
34.
Yamanaka H, Maehira F, Oshiro M, et al.:
A possible interaction of thioredoxin with VDUP1 in HeLa cells detected in a yeast two-hybrid system.
Biochem Biophys Res Commun.
2000; 271(3): 796–800. PubMed Abstract
| Publisher Full Text
36.
Patwari P, Chutkow WA, Cummings K, et al.:
Thioredoxin-independent regulation of metabolism by the alpha-arrestin proteins.
J Biol Chem.
2009; 284(37): 24996–25003. PubMed Abstract
| Publisher Full Text
| Free Full Text
37.
Patwari P, Emilsson V, Schadt EE, et al.:
The arrestin domain-containing 3 protein regulates body mass and energy expenditure.
Cell Metab.
2011; 14(5): 671–683. PubMed Abstract
| Publisher Full Text
| Free Full Text
39.
Park S, Jung Y, An SWA, et al.:
Dataset 1 in: RNAi targeting Caenorhabditis elegans. α-arrestins marginally affects lifespan.
F1000Research.
2017. Data Source
40.
Park S, Jung Y, An SWA, et al.:
Dataset 2 in: RNAi targeting Caenorhabditis elegans. α-arrestins marginally affects lifespan.
F1000Research.
2017. Data Source
41.
Park S, Jung Y, An SWA, et al.:
Dataset 3 in: RNAi targeting Caenorhabditis elegans. α-arrestins marginally affects lifespan.
F1000Research.
2017. Data Source
1
Department of Life Sciences, Pohang University of Science and Technology, Pohang, Gyeongbuk, 37673, South Korea 2
School of Interdisciplinary Bioscience and Bioengineering, Pohang University of Science and Technology, Pohang, Gyeongbuk, 37673, South Korea
This work was supported by the Korean Government (MSIP) through the National Research Foundation of Korea (NRF) [NRF-2016R1E1A1A01941152] funded to S-J.V.L.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Park S, Jung Y, An SWA et al. RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan [version 4; peer review: 3 approved]. F1000Research 2017, 6:1515 (https://doi.org/10.12688/f1000research.12337.4)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Biology of Aging
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
I appreciate the wish for the authors to move past this set of experiments and communicate their results to the field. Nevertheless, I must reiterate that, without testing the efficacy of their RNAi, their results cannot be interpreted, rather than be
... Continue reading
I appreciate the wish for the authors to move past this set of experiments and communicate their results to the field. Nevertheless, I must reiterate that, without testing the efficacy of their RNAi, their results cannot be interpreted, rather than be "interpreted with caution" as the authors suggest in the discussion.
Indeed, the authors themselves agree in their response to my previous review that "providing evidence for RNAi targeting arrd genes is a key for concluding that knockdown of arrd genes does not affect lifespan."
The authors suggest that performing qPCR on a new set of RNAi experiments would not demonstrate whether the knockdown worked on the worms followed for lifespan curves. While technically correct, I think it is fair to assume that their RNAi results are reproducible enough to warrant the assumption that whatever happens in a new set of RNAi experiments has happened in a previous one.
The authors bring forth a third reason for not testing the efficiency of their RNAi by claiming that their results are still useful to the field as they are. In fact, as I have suggested in my previous review, without qPCR testing of the efficacy of their RNAi, their results cannot be distinguished from the natural variation in lifespan of a control population of N2 worms, and are thus potentially meaningless.
In all honesty, I do not think that performing 24 qPCR reactions is an unreasonable request, especially when it is agreed that it would be "key for concluding that knockdown of arrd genes does not affect lifespan", which is the main point of the manuscript.
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Biology of Aging
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Author Response
(F1000Research Advisory Board Member)
08 Dec 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
08 Dec 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Alessandro Bitto,
We greatly appreciate your time and efforts for further valuable comments. We totally agree with your concern that we do not know the degree of RNAi knockdown
...
Continue readingDear Dr. Alessandro Bitto,
We greatly appreciate your time and efforts for further valuable comments. We totally agree with your concern that we do not know the degree of RNAi knockdown efficiency in worms except daf-16 RNAi, which was used as a positive control and showed expected lifespan phenotypes. Therefore, to precisely describe what we did, we changed “RNAi knockdown” to “feeding bacteria expressing dsRNA” throughout the manuscript. We sincerely thank you for fixing our misleading interpretation.
Dear Dr. Alessandro Bitto,
We greatly appreciate your time and efforts for further valuable comments. We totally agree with your concern that we do not know the degree of RNAi knockdown efficiency in worms except daf-16 RNAi, which was used as a positive control and showed expected lifespan phenotypes. Therefore, to precisely describe what we did, we changed “RNAi knockdown” to “feeding bacteria expressing dsRNA” throughout the manuscript. We sincerely thank you for fixing our misleading interpretation.
Competing Interests:No competing interests were disclosed.Close
Author Response
(F1000Research Advisory Board Member)
08 Dec 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
08 Dec 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Alessandro Bitto,
We greatly appreciate your time and efforts for further valuable comments. We totally agree with your concern that we do not know the degree of RNAi knockdown
...
Continue readingDear Dr. Alessandro Bitto,
We greatly appreciate your time and efforts for further valuable comments. We totally agree with your concern that we do not know the degree of RNAi knockdown efficiency in worms except daf-16 RNAi, which was used as a positive control and showed expected lifespan phenotypes. Therefore, to precisely describe what we did, we changed “RNAi knockdown” to “feeding bacteria expressing dsRNA” throughout the manuscript. We sincerely thank you for fixing our misleading interpretation.
Dear Dr. Alessandro Bitto,
We greatly appreciate your time and efforts for further valuable comments. We totally agree with your concern that we do not know the degree of RNAi knockdown efficiency in worms except daf-16 RNAi, which was used as a positive control and showed expected lifespan phenotypes. Therefore, to precisely describe what we did, we changed “RNAi knockdown” to “feeding bacteria expressing dsRNA” throughout the manuscript. We sincerely thank you for fixing our misleading interpretation.
Competing Interests:No competing interests were disclosed.Close
The authors took effort to address most of the concerns we raised in our initial review, but did not address at all our major concern, apart from a note in the discussion about the limitations of their results.
... Continue reading
The authors took effort to address most of the concerns we raised in our initial review, but did not address at all our major concern, apart from a note in the discussion about the limitations of their results.
Thus our major concern stands: the authors must provide evidence that RNAi knockdown is functioning for each of the genes tested before their lifespan results can be interpreted correctly. Until such evidence is provided, there is no way of determining whether knockdown of individual arrestins really has limited effect on lifespan.
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Biology of Aging
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Author Response
(F1000Research Advisory Board Member)
14 Nov 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
14 Nov 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Alessandro Bitto, We greatly appreciate your comments and respect your concerns on our paper. We totally agree with your point that providing evidence for RNAi targeting arrd genes is
...
Continue readingDear Dr. Alessandro Bitto, We greatly appreciate your comments and respect your concerns on our paper. We totally agree with your point that providing evidence for RNAi targeting arrd genes is a key for concluding that knockdown of arrd genes does not affect lifespan. Nevertheless, we think it is more beneficial for the research field and for our lab to publish our data as they are, instead of not publishing a paper or performing all the qRT-PCR experiments retrospectively because of the following reasons.
First, we emphasized the limitation and the pitfall of our data thoroughly and therefore the readers will be fully aware of that. Second, even if we perform the arrd RNAi experiments again and measure mRNA levels now, we will not be able to know whether the RNAi worked for the ones that we used for our lifespan screen. In addition, we do not have a plan on working on arrd genes any more. Third, with all these issues and limitations in mind, we think our negative data are still useful for the field. If some researchers want to do lifespan experiments for arrd genes in the future, they would rather use RNAi-sensitive strains and/or mutants by knowing our negative results. We think publishing these results is better for the field than keeping the data in our lab unpublished.
In any case, we sincerely appreciate your efforts for evaluating our manuscript.
Dear Dr. Alessandro Bitto, We greatly appreciate your comments and respect your concerns on our paper. We totally agree with your point that providing evidence for RNAi targeting arrd genes is a key for concluding that knockdown of arrd genes does not affect lifespan. Nevertheless, we think it is more beneficial for the research field and for our lab to publish our data as they are, instead of not publishing a paper or performing all the qRT-PCR experiments retrospectively because of the following reasons.
First, we emphasized the limitation and the pitfall of our data thoroughly and therefore the readers will be fully aware of that. Second, even if we perform the arrd RNAi experiments again and measure mRNA levels now, we will not be able to know whether the RNAi worked for the ones that we used for our lifespan screen. In addition, we do not have a plan on working on arrd genes any more. Third, with all these issues and limitations in mind, we think our negative data are still useful for the field. If some researchers want to do lifespan experiments for arrd genes in the future, they would rather use RNAi-sensitive strains and/or mutants by knowing our negative results. We think publishing these results is better for the field than keeping the data in our lab unpublished.
In any case, we sincerely appreciate your efforts for evaluating our manuscript.
Competing Interests:No competing interests were disclosed.Close
Author Response
(F1000Research Advisory Board Member)
14 Nov 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
14 Nov 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Alessandro Bitto, We greatly appreciate your comments and respect your concerns on our paper. We totally agree with your point that providing evidence for RNAi targeting arrd genes is
...
Continue readingDear Dr. Alessandro Bitto, We greatly appreciate your comments and respect your concerns on our paper. We totally agree with your point that providing evidence for RNAi targeting arrd genes is a key for concluding that knockdown of arrd genes does not affect lifespan. Nevertheless, we think it is more beneficial for the research field and for our lab to publish our data as they are, instead of not publishing a paper or performing all the qRT-PCR experiments retrospectively because of the following reasons.
First, we emphasized the limitation and the pitfall of our data thoroughly and therefore the readers will be fully aware of that. Second, even if we perform the arrd RNAi experiments again and measure mRNA levels now, we will not be able to know whether the RNAi worked for the ones that we used for our lifespan screen. In addition, we do not have a plan on working on arrd genes any more. Third, with all these issues and limitations in mind, we think our negative data are still useful for the field. If some researchers want to do lifespan experiments for arrd genes in the future, they would rather use RNAi-sensitive strains and/or mutants by knowing our negative results. We think publishing these results is better for the field than keeping the data in our lab unpublished.
In any case, we sincerely appreciate your efforts for evaluating our manuscript.
Dear Dr. Alessandro Bitto, We greatly appreciate your comments and respect your concerns on our paper. We totally agree with your point that providing evidence for RNAi targeting arrd genes is a key for concluding that knockdown of arrd genes does not affect lifespan. Nevertheless, we think it is more beneficial for the research field and for our lab to publish our data as they are, instead of not publishing a paper or performing all the qRT-PCR experiments retrospectively because of the following reasons.
First, we emphasized the limitation and the pitfall of our data thoroughly and therefore the readers will be fully aware of that. Second, even if we perform the arrd RNAi experiments again and measure mRNA levels now, we will not be able to know whether the RNAi worked for the ones that we used for our lifespan screen. In addition, we do not have a plan on working on arrd genes any more. Third, with all these issues and limitations in mind, we think our negative data are still useful for the field. If some researchers want to do lifespan experiments for arrd genes in the future, they would rather use RNAi-sensitive strains and/or mutants by knowing our negative results. We think publishing these results is better for the field than keeping the data in our lab unpublished.
In any case, we sincerely appreciate your efforts for evaluating our manuscript.
Competing Interests:No competing interests were disclosed.Close
In this paper, Park et al. knocked down 24 out of 29 alpha-arrestin genes in C. elegans and found that none of them produced a sizable effect on the lifespans of the wild type or the long-lived daf-2 worms. The
... Continue reading
In this paper, Park et al. knocked down 24 out of 29 alpha-arrestin genes in C. elegans and found that none of them produced a sizable effect on the lifespans of the wild type or the long-lived daf-2 worms. The methods used in this study were sound, and the data quality were up to standard with hundreds of worms assayed for RNAi of each alpha-arrestin gene. I think the work is honest, and the negative results in this paper are reliable.
Concerns:
(1) I agree with the previous 2 reviewers that it is strange to leave out 5 members of the C. elegans alpha-arrestin genes.
(2) I think it is fair to say that none of the 24 genes extended lifespan when knocked down. The statistically significant 10% increase of wild-type lifespan by arrd-3 RNAi is negligible, given the variation of the lifespan assay. The relatively small p-value (0.0045) is blessed by the large sample size (n = 240). I wonder what the p-values would be if the two independent lifespan experiments had not been pooled.
(3) I would like the authors to report how many worms were censored.
(4) I wonder whether the authors had observed any noticeable phenotypes from their RNAi experiments, in particular, egg-laying defects. Strong egg-laying defects are easy to detect. G-protein signaling and beta-arrestin regulate the egg-laying behavior.
Is the work clearly and accurately presented and does it cite the current literature?
Yes
Is the study design appropriate and is the work technically sound?
Yes
Are sufficient details of methods and analysis provided to allow replication by others?
Yes
If applicable, is the statistical analysis and its interpretation appropriate?
I cannot comment. A qualified statistician is required.
Are all the source data underlying the results available to ensure full reproducibility?
Partly
Are the conclusions drawn adequately supported by the results?
Yes
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: C. elegans aging
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Author Response
(F1000Research Advisory Board Member)
14 Nov 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
14 Nov 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Meng-Qiu Dong, We sincerely thank you for your valuable comments on our manuscript. We admit the limitations of our research and revised our manuscript to downplay our results so
...
Continue readingDear Dr. Meng-Qiu Dong, We sincerely thank you for your valuable comments on our manuscript. We admit the limitations of our research and revised our manuscript to downplay our results so as not to confuse the readers. We addressed your concerns and revised our manuscript as described below.
In this paper, Park et al. knocked down 24 out of 29 alpha-arrestin genes in C. elegans and found that none of them produced a sizable effect on the lifespans of the wild type or the long-lived daf-2 worms. The methods used in this study were sound, and the data quality were up to standard with hundreds of worms assayed for RNAi of each alpha-arrestin gene. I think the work is honest, and the negative results in this paper are reliable.
Concerns: (1) I agree with the previous 2 reviewers that it is strange to leave out 5 members of the C. elegans alpha-arrestin genes.
> We appreciate your comments. We leave out arrd-20 and arrd-21 because they are predicted to be pseudogenes (Wormbase, version WS259) that are expected not to encode functional proteins. In addition, we did not test arrd-12, arrd-22 or arrd-27 because we were not able to obtain the corresponding RNAi clones during our cloning. We addressed this issue in Methods and discussed potential lifespan-regulatory roles of those genes in Discussion as follows, In Methods, RNAi clones: “RNAi clone for arrd-12, arrd-22, or arrd-27 was not generated because we were not able to obtain transformed E. coli (HT115) colonies using arrd-12 or arrd-22 infusion reaction products, or the PCR product for arrd-27 genomic or complementary DNA.” In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
(2) I think it is fair to say that none of the 24 genes extended lifespan when knocked down. The statistically significant 10% increase of wild-type lifespan by arrd-3 RNAi is negligible, given the variation of the lifespan assay. The relatively small p-value (0.0045) is blessed by the large sample size (n = 240). I wonder what the p-values would be if the two independent lifespan experiments had not been pooled.
> We appreciate your valuable comments. We agree with you that the extent of lifespan changes by several arrd RNAi clones was small but statistically significant because of the large sample size. Even if the lifespan data are not pooled, many independent lifespan results for arrd RNAi clones were again statistically significant. In either case, pooled or not pooled, we think the main point of our data indicates little effects of arrd RNAi on lifespan.
(3) I would like the authors to report how many worms were censored.
> We appreciate your comments. We provided raw lifespan data that contain the number of censored worms on each day of counting in Dataset1. One can also easily know the total number of censored worms by the difference between the number of animals that died and the total number of worms in Supplementary Table S2.
(4) I wonder whether the authors had observed any noticeable phenotypes from their RNAi experiments, in particular, egg-laying defects. Strong egg-laying defects are easy to detect. G-protein signaling and beta-arrestin regulate the egg-laying behavior.
> We appreciate your comments. Because we focused on the lifespan phenotype of arrd RNAi, unfortunately we did not pay attention to other physiological phenotypes and might have missed egg-laying phenotypes caused by some arrd RNAi clones. It will be an important point for future research.
Dear Dr. Meng-Qiu Dong, We sincerely thank you for your valuable comments on our manuscript. We admit the limitations of our research and revised our manuscript to downplay our results so as not to confuse the readers. We addressed your concerns and revised our manuscript as described below.
In this paper, Park et al. knocked down 24 out of 29 alpha-arrestin genes in C. elegans and found that none of them produced a sizable effect on the lifespans of the wild type or the long-lived daf-2 worms. The methods used in this study were sound, and the data quality were up to standard with hundreds of worms assayed for RNAi of each alpha-arrestin gene. I think the work is honest, and the negative results in this paper are reliable.
Concerns: (1) I agree with the previous 2 reviewers that it is strange to leave out 5 members of the C. elegans alpha-arrestin genes.
> We appreciate your comments. We leave out arrd-20 and arrd-21 because they are predicted to be pseudogenes (Wormbase, version WS259) that are expected not to encode functional proteins. In addition, we did not test arrd-12, arrd-22 or arrd-27 because we were not able to obtain the corresponding RNAi clones during our cloning. We addressed this issue in Methods and discussed potential lifespan-regulatory roles of those genes in Discussion as follows, In Methods, RNAi clones: “RNAi clone for arrd-12, arrd-22, or arrd-27 was not generated because we were not able to obtain transformed E. coli (HT115) colonies using arrd-12 or arrd-22 infusion reaction products, or the PCR product for arrd-27 genomic or complementary DNA.” In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
(2) I think it is fair to say that none of the 24 genes extended lifespan when knocked down. The statistically significant 10% increase of wild-type lifespan by arrd-3 RNAi is negligible, given the variation of the lifespan assay. The relatively small p-value (0.0045) is blessed by the large sample size (n = 240). I wonder what the p-values would be if the two independent lifespan experiments had not been pooled.
> We appreciate your valuable comments. We agree with you that the extent of lifespan changes by several arrd RNAi clones was small but statistically significant because of the large sample size. Even if the lifespan data are not pooled, many independent lifespan results for arrd RNAi clones were again statistically significant. In either case, pooled or not pooled, we think the main point of our data indicates little effects of arrd RNAi on lifespan.
(3) I would like the authors to report how many worms were censored.
> We appreciate your comments. We provided raw lifespan data that contain the number of censored worms on each day of counting in Dataset1. One can also easily know the total number of censored worms by the difference between the number of animals that died and the total number of worms in Supplementary Table S2.
(4) I wonder whether the authors had observed any noticeable phenotypes from their RNAi experiments, in particular, egg-laying defects. Strong egg-laying defects are easy to detect. G-protein signaling and beta-arrestin regulate the egg-laying behavior.
> We appreciate your comments. Because we focused on the lifespan phenotype of arrd RNAi, unfortunately we did not pay attention to other physiological phenotypes and might have missed egg-laying phenotypes caused by some arrd RNAi clones. It will be an important point for future research.
Competing Interests:No competing interests were disclosed.Close
Author Response
(F1000Research Advisory Board Member)
14 Nov 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
14 Nov 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Meng-Qiu Dong, We sincerely thank you for your valuable comments on our manuscript. We admit the limitations of our research and revised our manuscript to downplay our results so
...
Continue readingDear Dr. Meng-Qiu Dong, We sincerely thank you for your valuable comments on our manuscript. We admit the limitations of our research and revised our manuscript to downplay our results so as not to confuse the readers. We addressed your concerns and revised our manuscript as described below.
In this paper, Park et al. knocked down 24 out of 29 alpha-arrestin genes in C. elegans and found that none of them produced a sizable effect on the lifespans of the wild type or the long-lived daf-2 worms. The methods used in this study were sound, and the data quality were up to standard with hundreds of worms assayed for RNAi of each alpha-arrestin gene. I think the work is honest, and the negative results in this paper are reliable.
Concerns: (1) I agree with the previous 2 reviewers that it is strange to leave out 5 members of the C. elegans alpha-arrestin genes.
> We appreciate your comments. We leave out arrd-20 and arrd-21 because they are predicted to be pseudogenes (Wormbase, version WS259) that are expected not to encode functional proteins. In addition, we did not test arrd-12, arrd-22 or arrd-27 because we were not able to obtain the corresponding RNAi clones during our cloning. We addressed this issue in Methods and discussed potential lifespan-regulatory roles of those genes in Discussion as follows, In Methods, RNAi clones: “RNAi clone for arrd-12, arrd-22, or arrd-27 was not generated because we were not able to obtain transformed E. coli (HT115) colonies using arrd-12 or arrd-22 infusion reaction products, or the PCR product for arrd-27 genomic or complementary DNA.” In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
(2) I think it is fair to say that none of the 24 genes extended lifespan when knocked down. The statistically significant 10% increase of wild-type lifespan by arrd-3 RNAi is negligible, given the variation of the lifespan assay. The relatively small p-value (0.0045) is blessed by the large sample size (n = 240). I wonder what the p-values would be if the two independent lifespan experiments had not been pooled.
> We appreciate your valuable comments. We agree with you that the extent of lifespan changes by several arrd RNAi clones was small but statistically significant because of the large sample size. Even if the lifespan data are not pooled, many independent lifespan results for arrd RNAi clones were again statistically significant. In either case, pooled or not pooled, we think the main point of our data indicates little effects of arrd RNAi on lifespan.
(3) I would like the authors to report how many worms were censored.
> We appreciate your comments. We provided raw lifespan data that contain the number of censored worms on each day of counting in Dataset1. One can also easily know the total number of censored worms by the difference between the number of animals that died and the total number of worms in Supplementary Table S2.
(4) I wonder whether the authors had observed any noticeable phenotypes from their RNAi experiments, in particular, egg-laying defects. Strong egg-laying defects are easy to detect. G-protein signaling and beta-arrestin regulate the egg-laying behavior.
> We appreciate your comments. Because we focused on the lifespan phenotype of arrd RNAi, unfortunately we did not pay attention to other physiological phenotypes and might have missed egg-laying phenotypes caused by some arrd RNAi clones. It will be an important point for future research.
Dear Dr. Meng-Qiu Dong, We sincerely thank you for your valuable comments on our manuscript. We admit the limitations of our research and revised our manuscript to downplay our results so as not to confuse the readers. We addressed your concerns and revised our manuscript as described below.
In this paper, Park et al. knocked down 24 out of 29 alpha-arrestin genes in C. elegans and found that none of them produced a sizable effect on the lifespans of the wild type or the long-lived daf-2 worms. The methods used in this study were sound, and the data quality were up to standard with hundreds of worms assayed for RNAi of each alpha-arrestin gene. I think the work is honest, and the negative results in this paper are reliable.
Concerns: (1) I agree with the previous 2 reviewers that it is strange to leave out 5 members of the C. elegans alpha-arrestin genes.
> We appreciate your comments. We leave out arrd-20 and arrd-21 because they are predicted to be pseudogenes (Wormbase, version WS259) that are expected not to encode functional proteins. In addition, we did not test arrd-12, arrd-22 or arrd-27 because we were not able to obtain the corresponding RNAi clones during our cloning. We addressed this issue in Methods and discussed potential lifespan-regulatory roles of those genes in Discussion as follows, In Methods, RNAi clones: “RNAi clone for arrd-12, arrd-22, or arrd-27 was not generated because we were not able to obtain transformed E. coli (HT115) colonies using arrd-12 or arrd-22 infusion reaction products, or the PCR product for arrd-27 genomic or complementary DNA.” In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
(2) I think it is fair to say that none of the 24 genes extended lifespan when knocked down. The statistically significant 10% increase of wild-type lifespan by arrd-3 RNAi is negligible, given the variation of the lifespan assay. The relatively small p-value (0.0045) is blessed by the large sample size (n = 240). I wonder what the p-values would be if the two independent lifespan experiments had not been pooled.
> We appreciate your valuable comments. We agree with you that the extent of lifespan changes by several arrd RNAi clones was small but statistically significant because of the large sample size. Even if the lifespan data are not pooled, many independent lifespan results for arrd RNAi clones were again statistically significant. In either case, pooled or not pooled, we think the main point of our data indicates little effects of arrd RNAi on lifespan.
(3) I would like the authors to report how many worms were censored.
> We appreciate your comments. We provided raw lifespan data that contain the number of censored worms on each day of counting in Dataset1. One can also easily know the total number of censored worms by the difference between the number of animals that died and the total number of worms in Supplementary Table S2.
(4) I wonder whether the authors had observed any noticeable phenotypes from their RNAi experiments, in particular, egg-laying defects. Strong egg-laying defects are easy to detect. G-protein signaling and beta-arrestin regulate the egg-laying behavior.
> We appreciate your comments. Because we focused on the lifespan phenotype of arrd RNAi, unfortunately we did not pay attention to other physiological phenotypes and might have missed egg-laying phenotypes caused by some arrd RNAi clones. It will be an important point for future research.
Competing Interests:No competing interests were disclosed.Close
NOTE: it is important to ensure the information in square brackets after the title is included in this citation.
Reviewer Report20 Oct 2017
Roger Pocock,
Development and Stem Cells Program, Monash Biomedicine Discovery Institute, Department of Anatomy and Developmental Biology, Monash University, Melbourne, Vic, Australia
The authors have partially addressed my concerns. Below I suggest further changes.
Change title from 'RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan' to 'RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan'
... Continue reading
The authors have partially addressed my concerns. Below I suggest further changes.
Change title from 'RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan' to 'RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan'
Also change from 'small or no effects' to 'little effect' throughout the text.
The authors state that they did not study arrd-12, 22 and 27 because of 'unknown technical problems'. It would be helpful for the field to be more specific here. Could they not be amplified?
In the results paragraph the authors state that 'knockdown of each of individual α-arrestin genes tended to slightly reduce lifespan in wild-type or in daf-2(-) mutants'
This is a little misleading. Change to 'knockdown of specific α-arrestin genes caused a minor reduction of lifespan in wild-type or in daf-2(-) mutants'
Change from 'In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR' to 'In addition, as we did not test whether RNAi targeting of each α-arrestin gene was effective by using quantitative RT-PCR'
Competing Interests: No competing interests were disclosed.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Author Response
(F1000Research Advisory Board Member)
14 Nov 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
14 Nov 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Roger Pocock, We greatly appreciate your comments on our manuscript. We further addressed your concerns and revised our manuscript as described below.
The authors have partially addressed my concerns. Below
...
Continue readingDear Dr. Roger Pocock, We greatly appreciate your comments on our manuscript. We further addressed your concerns and revised our manuscript as described below.
The authors have partially addressed my concerns. Below I suggest further changes. 1. Change title from 'RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan' to 'RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan'
> We appreciate your comment on the title. We changed the title as follows, Title: RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan
2. Also change from 'small or no effects' to 'little effect' throughout the text.
> We also changed the ‘small or no effects’ to ‘little effect’ in the main text as follows, In Abstract: “We found that knocking down each of 24 out of total 29 C. elegans α-arrestins had little effect on lifespan.” In Introduction: “We found that knocking down each of the α-arrestins had little effect on the lifespan of wild-type and daf-2 mutants.” In Discussion: “Here we showed that RNAi knockdown of individual C. elegans α-arrestins had little effect on lifespan in wild-type or daf-2 mutants.” In Figure legend: “Figure 1. RNAi targeting each of Caenorhabditis elegans α-arrestins had little effect on lifespan.”
3. The authors state that they did not study arrd-12, 22 and 27 because of 'unknown technical problems'. It would be helpful for the field to be more specific here. Could they not be amplified?
> We appreciate your comments. We were not able to obtain transformed E. coli HT115 with infusion reaction products for arrd-12 or arrd-22 RNAi clones, or the PCR product for arrd-27 genomic or complementary DNA. We specified the ‘unknown technical problems’ by mentioning these details in the main text as follows, In Methods, RNAi clones: “RNAi clone for arrd-12, arrd-22, or arrd-27 was not generated because we were not able to obtain transformed E. coli (HT115) colonies using arrd-12 or arrd-22 infusion reaction products, or the PCR product for arrd-27 genomic or complementary DNA.”
4. In the results paragraph the authors state that 'knockdown of each of individual α-arrestin genes tended to slightly reduce lifespan in wild-type or in daf-2(-) mutants' This is a little misleading. Change to 'knockdown of specific α-arrestin genes caused a minor reduction of lifespan in wild-type or in daf-2(-) mutants'
> We appreciate your comments. We changed the sentence as follows, In Results: “We found that knockdown of specific α-arrestin genes caused a minor reduction of lifespan in wild-type or in daf-2(-) mutants (Figure 1B, Figure S1B-U, Table S2).”
5. Change from 'In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR' to 'In addition, as we did not test whether RNAi targeting of each α-arrestin gene was effective by using quantitative RT-PCR'
> We appreciate your comments. We changed the sentence as suggested. In Discussion: “In addition, as we did not test whether RNAi targeting each α-arrestin gene was effective by using quantitative RT-PCR”
Dear Dr. Roger Pocock, We greatly appreciate your comments on our manuscript. We further addressed your concerns and revised our manuscript as described below.
The authors have partially addressed my concerns. Below I suggest further changes. 1. Change title from 'RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan' to 'RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan'
> We appreciate your comment on the title. We changed the title as follows, Title: RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan
2. Also change from 'small or no effects' to 'little effect' throughout the text.
> We also changed the ‘small or no effects’ to ‘little effect’ in the main text as follows, In Abstract: “We found that knocking down each of 24 out of total 29 C. elegans α-arrestins had little effect on lifespan.” In Introduction: “We found that knocking down each of the α-arrestins had little effect on the lifespan of wild-type and daf-2 mutants.” In Discussion: “Here we showed that RNAi knockdown of individual C. elegans α-arrestins had little effect on lifespan in wild-type or daf-2 mutants.” In Figure legend: “Figure 1. RNAi targeting each of Caenorhabditis elegans α-arrestins had little effect on lifespan.”
3. The authors state that they did not study arrd-12, 22 and 27 because of 'unknown technical problems'. It would be helpful for the field to be more specific here. Could they not be amplified?
> We appreciate your comments. We were not able to obtain transformed E. coli HT115 with infusion reaction products for arrd-12 or arrd-22 RNAi clones, or the PCR product for arrd-27 genomic or complementary DNA. We specified the ‘unknown technical problems’ by mentioning these details in the main text as follows, In Methods, RNAi clones: “RNAi clone for arrd-12, arrd-22, or arrd-27 was not generated because we were not able to obtain transformed E. coli (HT115) colonies using arrd-12 or arrd-22 infusion reaction products, or the PCR product for arrd-27 genomic or complementary DNA.”
4. In the results paragraph the authors state that 'knockdown of each of individual α-arrestin genes tended to slightly reduce lifespan in wild-type or in daf-2(-) mutants' This is a little misleading. Change to 'knockdown of specific α-arrestin genes caused a minor reduction of lifespan in wild-type or in daf-2(-) mutants'
> We appreciate your comments. We changed the sentence as follows, In Results: “We found that knockdown of specific α-arrestin genes caused a minor reduction of lifespan in wild-type or in daf-2(-) mutants (Figure 1B, Figure S1B-U, Table S2).”
5. Change from 'In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR' to 'In addition, as we did not test whether RNAi targeting of each α-arrestin gene was effective by using quantitative RT-PCR'
> We appreciate your comments. We changed the sentence as suggested. In Discussion: “In addition, as we did not test whether RNAi targeting each α-arrestin gene was effective by using quantitative RT-PCR”
Competing Interests:No competing interests were disclosed.Close
Author Response
(F1000Research Advisory Board Member)
14 Nov 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
14 Nov 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Roger Pocock, We greatly appreciate your comments on our manuscript. We further addressed your concerns and revised our manuscript as described below.
The authors have partially addressed my concerns. Below
...
Continue readingDear Dr. Roger Pocock, We greatly appreciate your comments on our manuscript. We further addressed your concerns and revised our manuscript as described below.
The authors have partially addressed my concerns. Below I suggest further changes. 1. Change title from 'RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan' to 'RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan'
> We appreciate your comment on the title. We changed the title as follows, Title: RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan
2. Also change from 'small or no effects' to 'little effect' throughout the text.
> We also changed the ‘small or no effects’ to ‘little effect’ in the main text as follows, In Abstract: “We found that knocking down each of 24 out of total 29 C. elegans α-arrestins had little effect on lifespan.” In Introduction: “We found that knocking down each of the α-arrestins had little effect on the lifespan of wild-type and daf-2 mutants.” In Discussion: “Here we showed that RNAi knockdown of individual C. elegans α-arrestins had little effect on lifespan in wild-type or daf-2 mutants.” In Figure legend: “Figure 1. RNAi targeting each of Caenorhabditis elegans α-arrestins had little effect on lifespan.”
3. The authors state that they did not study arrd-12, 22 and 27 because of 'unknown technical problems'. It would be helpful for the field to be more specific here. Could they not be amplified?
> We appreciate your comments. We were not able to obtain transformed E. coli HT115 with infusion reaction products for arrd-12 or arrd-22 RNAi clones, or the PCR product for arrd-27 genomic or complementary DNA. We specified the ‘unknown technical problems’ by mentioning these details in the main text as follows, In Methods, RNAi clones: “RNAi clone for arrd-12, arrd-22, or arrd-27 was not generated because we were not able to obtain transformed E. coli (HT115) colonies using arrd-12 or arrd-22 infusion reaction products, or the PCR product for arrd-27 genomic or complementary DNA.”
4. In the results paragraph the authors state that 'knockdown of each of individual α-arrestin genes tended to slightly reduce lifespan in wild-type or in daf-2(-) mutants' This is a little misleading. Change to 'knockdown of specific α-arrestin genes caused a minor reduction of lifespan in wild-type or in daf-2(-) mutants'
> We appreciate your comments. We changed the sentence as follows, In Results: “We found that knockdown of specific α-arrestin genes caused a minor reduction of lifespan in wild-type or in daf-2(-) mutants (Figure 1B, Figure S1B-U, Table S2).”
5. Change from 'In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR' to 'In addition, as we did not test whether RNAi targeting of each α-arrestin gene was effective by using quantitative RT-PCR'
> We appreciate your comments. We changed the sentence as suggested. In Discussion: “In addition, as we did not test whether RNAi targeting each α-arrestin gene was effective by using quantitative RT-PCR”
Dear Dr. Roger Pocock, We greatly appreciate your comments on our manuscript. We further addressed your concerns and revised our manuscript as described below.
The authors have partially addressed my concerns. Below I suggest further changes. 1. Change title from 'RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan' to 'RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan'
> We appreciate your comment on the title. We changed the title as follows, Title: RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan
2. Also change from 'small or no effects' to 'little effect' throughout the text.
> We also changed the ‘small or no effects’ to ‘little effect’ in the main text as follows, In Abstract: “We found that knocking down each of 24 out of total 29 C. elegans α-arrestins had little effect on lifespan.” In Introduction: “We found that knocking down each of the α-arrestins had little effect on the lifespan of wild-type and daf-2 mutants.” In Discussion: “Here we showed that RNAi knockdown of individual C. elegans α-arrestins had little effect on lifespan in wild-type or daf-2 mutants.” In Figure legend: “Figure 1. RNAi targeting each of Caenorhabditis elegans α-arrestins had little effect on lifespan.”
3. The authors state that they did not study arrd-12, 22 and 27 because of 'unknown technical problems'. It would be helpful for the field to be more specific here. Could they not be amplified?
> We appreciate your comments. We were not able to obtain transformed E. coli HT115 with infusion reaction products for arrd-12 or arrd-22 RNAi clones, or the PCR product for arrd-27 genomic or complementary DNA. We specified the ‘unknown technical problems’ by mentioning these details in the main text as follows, In Methods, RNAi clones: “RNAi clone for arrd-12, arrd-22, or arrd-27 was not generated because we were not able to obtain transformed E. coli (HT115) colonies using arrd-12 or arrd-22 infusion reaction products, or the PCR product for arrd-27 genomic or complementary DNA.”
4. In the results paragraph the authors state that 'knockdown of each of individual α-arrestin genes tended to slightly reduce lifespan in wild-type or in daf-2(-) mutants' This is a little misleading. Change to 'knockdown of specific α-arrestin genes caused a minor reduction of lifespan in wild-type or in daf-2(-) mutants'
> We appreciate your comments. We changed the sentence as follows, In Results: “We found that knockdown of specific α-arrestin genes caused a minor reduction of lifespan in wild-type or in daf-2(-) mutants (Figure 1B, Figure S1B-U, Table S2).”
5. Change from 'In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR' to 'In addition, as we did not test whether RNAi targeting of each α-arrestin gene was effective by using quantitative RT-PCR'
> We appreciate your comments. We changed the sentence as suggested. In Discussion: “In addition, as we did not test whether RNAi targeting each α-arrestin gene was effective by using quantitative RT-PCR”
Competing Interests:No competing interests were disclosed.Close
NOTE: it is important to ensure the information in square brackets after the title is included in this citation.
Reviewer Report06 Sep 2017
Roger Pocock,
Development and Stem Cells Program, Monash Biomedicine Discovery Institute, Department of Anatomy and Developmental Biology, Monash University, Melbourne, Vic, Australia
In this manuscript Park et al. performed RNAi knockdown of 24 of the 29 alpha-arrestin genes in C. elegans to ask whether they are important for lifespan regulation in wild type and daf-2 mutant animals. Their data shows that, in
... Continue reading
In this manuscript Park et al. performed RNAi knockdown of 24 of the 29 alpha-arrestin genes in C. elegans to ask whether they are important for lifespan regulation in wild type and daf-2 mutant animals. Their data shows that, in their experimental conditions, most alpha-arrestin genes are individually not important for regulating lifespan.
Concerns:
One major issue with the study is whether RNAi knockdown of the alpha-arrestin genes is working. As some alpha-arrestins are expressed in the nervous system, which is refractory to RNAi, this is a concern. To determine whether the RNAi knockdown is working then Q-RTPCR should be performed. However, this is not a small task for 24 genes, therefore, further experiments using sensitized genetic backgrounds for RNAi or elevated temperatures may tease out phenotypes for some alpha-arrestin genes. This could particularly be performed for the alpha-arrestin genes that when knocked down engender a slight effect on lifespan.
In parallel to this, the authors could align the alpha-arrestin gene sequences to ask if there is a common sequence that could be used by RNAi targeting to knockdown multiple alpha-arrestin genes simultaneously.
The authors knockdown of 24 of the 29 alpha-arrestin genes. I wonder why they didn’t generate RNAi clones for the remaining 5, especially since they did generate 3 other constructs for their analysis. Potentially, the important alpha-arrestin genes is amongst those not examined.
The authors did not report any other behavioural or morphological phenotypes observed by alpha-arrestin gene knockdown. Any information on observed phenotypes would be of interest to the research community.
Is the work clearly and accurately presented and does it cite the current literature?
Yes
Is the study design appropriate and is the work technically sound?
Partly
Are sufficient details of methods and analysis provided to allow replication by others?
Yes
If applicable, is the statistical analysis and its interpretation appropriate?
Partly
Are all the source data underlying the results available to ensure full reproducibility?
Yes
Are the conclusions drawn adequately supported by the results?
Partly
Competing Interests: No competing interests were disclosed.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Author Response
(F1000Research Advisory Board Member)
14 Nov 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
14 Nov 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Roger Pocock,
We sincerely appreciate your valuable comments on our manuscript. We agree with all your points and revised our manuscript to describe the limitations of this study.
...
Continue readingDear Dr. Roger Pocock,
We sincerely appreciate your valuable comments on our manuscript. We agree with all your points and revised our manuscript to describe the limitations of this study. We would like to mention that the main motivation for publishing this manuscript is to provide our negative results to researchers who are interested in characterizing α-arrestins in C. elegans. Our point-by-point responses are described below. We hope you find our revised manuscript is suitable for publication in F1000Research.
In this manuscript Park et al. performed RNAi knockdown of 24 of the 29 alpha-arrestin genes in C. elegans to ask whether they are important for lifespan regulation in wild type and daf-2 mutant animals. Their data shows that, in their experimental conditions, most alpha-arrestin genes are individually not important for regulating lifespan.
Concerns:
One major issue with the study is whether RNAi knockdown of the alpha-arrestin genes is working. As some alpha-arrestins are expressed in the nervous system, which is refractory to RNAi, this is a concern. To determine whether the RNAi knockdown is working then Q-RTPCR should be performed. However, this is not a small task for 24 genes, therefore, further experiments using sensitized genetic backgrounds for RNAi or elevated temperatures may tease out phenotypes for some alpha-arrestin genes. This could particularly be performed for the alpha-arrestin genes that when knocked down engender a slight effect on lifespan.
> We agree with the reviewer’s comment that the RNAi knockdown efficiency is a major concern in interpreting our screen results. It would have been great if we had performed qRT-PCR analysis. However, as the reviewers pointed out, this is not a small task. We currently do not have any plan for following up this project and want to inform other researchers of our screen data. Therefore, we further emphasized the pitfall of our results in the Discussion as follows,
In Discussion: “The lifespan-regulatory functions of α-arrestins remain largely unexplored at the organism level. Here we showed that RNAi knockdown of individual C. elegans α-arrestins had small or no effects on lifespan in wild-type or daf-2 mutants. Our study has limitations that need to be considered for interpretation. First, RNAi targeting some α-arrestins might be insufficient for causing strong lifespan phenotypes. This may be because many of C. elegans α-arrestins are predicted to be expressed in neurons7, 9, 24-26, which are refractory to RNAi27-29. Lifespan assays using RNAi-hypersensitive mutants, including rrf-3(-) and eri-1(-) animals29-31, treated with α-arrestin RNAi, or using α-arrestin null mutants will help address this issue. In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR, our negative data should be interpreted with caution.”
In parallel to this, the authors could align the alpha-arrestin gene sequences to ask if there is a common sequence that could be used by RNAi targeting to knockdown multiple alpha-arrestin genes simultaneously.
> We appreciate the reviewer’s comment. The α-arrestin family members share arrestin-domains (O. Hobert, Wormbook, 2013). Thus, it will be intriguing if we could examine whether targeting arrestin domain for RNAi to knock down multiple arrd genes simultaneously and if multiple arrd gene knock down has effects on lifespan. We modified our manuscript to address this issue in Discussion as follows,
In Discussion: “Second, C. elegans α-arrestins may have functional redundancy considering the large number of the α-arrestin family members in C. elegans and their sequence similarity1, 7, which may obscure examining the functional importance of each α-arrestin. In addition, some α-arrestins may mostly function by modulating their interacting proteins6. In this case, genetic inhibition of α-arrestins may rather subtly affect the functions of their interacting partners that directly regulate physiology, such as aging and longevity, causing weak or no phenotypes. Thus, it will be interesting to test the effects of simultaneous inhibition of α-arrestins possibly through targeting the arrestin domain, and to identify and to functionally characterize proteins that bind C. elegans α-arrestins.”
The authors knockdown of 24 of the 29 alpha-arrestin genes. I wonder why they didn’t generate RNAi clones for the remaining 5, especially since they did generate 3 other constructs for their analysis. Potentially, the important alpha-arrestin genes is amongst those not examined.
> We appreciate the reviewers’ comment. We aimed at obtaining all the α arrestin RNAi clones that are not commercially available, except two predicted pseudogenes, arrd-20 and arrd-21 (Wormbase, version WS259). We successfully constructed three RNAi clones targeting arrd-11, arrd-17 and arrd-26, but could not obtain the other three RNAi clones, targeting arrd-12, arrd-22 and arrd-27. We addressed this issue in Methods for RNAi clones and in Discussion as follows,
In Methods, RNAi clones: “RNAi clones for arrd-12, arrd-22, and arrd-27 were not generated because of unknown technical problems and were not included in the lifespan screen. arrd-20 and arrd-21 are predicted to be pseudogenes (Wormbase, version WS259) and were excluded from our screen.”
In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
The authors did not report any other behavioural or morphological phenotypes observed by alpha-arrestin gene knockdown. Any information on observed phenotypes would be of interest to the research community.
> We thank the reviewer for the valuable comment. One noticeable phenotype we observed was that the eggs laid by worms fed with ttm-2 RNAi bacteria hatched more frequently than control eggs on FUdR-treated plates, although we did not quantify the results. We described this issue in Methods as follows,
In Methods, RNAi screen using lifespan assays: “Eggs laid by ttm-2 RNAi-treated worms hatched more frequently than control eggs on FUdR-containing plates in two independent lifespan experiments.”
Dear Dr. Roger Pocock,
We sincerely appreciate your valuable comments on our manuscript. We agree with all your points and revised our manuscript to describe the limitations of this study. We would like to mention that the main motivation for publishing this manuscript is to provide our negative results to researchers who are interested in characterizing α-arrestins in C. elegans. Our point-by-point responses are described below. We hope you find our revised manuscript is suitable for publication in F1000Research.
In this manuscript Park et al. performed RNAi knockdown of 24 of the 29 alpha-arrestin genes in C. elegans to ask whether they are important for lifespan regulation in wild type and daf-2 mutant animals. Their data shows that, in their experimental conditions, most alpha-arrestin genes are individually not important for regulating lifespan.
Concerns:
One major issue with the study is whether RNAi knockdown of the alpha-arrestin genes is working. As some alpha-arrestins are expressed in the nervous system, which is refractory to RNAi, this is a concern. To determine whether the RNAi knockdown is working then Q-RTPCR should be performed. However, this is not a small task for 24 genes, therefore, further experiments using sensitized genetic backgrounds for RNAi or elevated temperatures may tease out phenotypes for some alpha-arrestin genes. This could particularly be performed for the alpha-arrestin genes that when knocked down engender a slight effect on lifespan.
> We agree with the reviewer’s comment that the RNAi knockdown efficiency is a major concern in interpreting our screen results. It would have been great if we had performed qRT-PCR analysis. However, as the reviewers pointed out, this is not a small task. We currently do not have any plan for following up this project and want to inform other researchers of our screen data. Therefore, we further emphasized the pitfall of our results in the Discussion as follows,
In Discussion: “The lifespan-regulatory functions of α-arrestins remain largely unexplored at the organism level. Here we showed that RNAi knockdown of individual C. elegans α-arrestins had small or no effects on lifespan in wild-type or daf-2 mutants. Our study has limitations that need to be considered for interpretation. First, RNAi targeting some α-arrestins might be insufficient for causing strong lifespan phenotypes. This may be because many of C. elegans α-arrestins are predicted to be expressed in neurons7, 9, 24-26, which are refractory to RNAi27-29. Lifespan assays using RNAi-hypersensitive mutants, including rrf-3(-) and eri-1(-) animals29-31, treated with α-arrestin RNAi, or using α-arrestin null mutants will help address this issue. In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR, our negative data should be interpreted with caution.”
In parallel to this, the authors could align the alpha-arrestin gene sequences to ask if there is a common sequence that could be used by RNAi targeting to knockdown multiple alpha-arrestin genes simultaneously.
> We appreciate the reviewer’s comment. The α-arrestin family members share arrestin-domains (O. Hobert, Wormbook, 2013). Thus, it will be intriguing if we could examine whether targeting arrestin domain for RNAi to knock down multiple arrd genes simultaneously and if multiple arrd gene knock down has effects on lifespan. We modified our manuscript to address this issue in Discussion as follows,
In Discussion: “Second, C. elegans α-arrestins may have functional redundancy considering the large number of the α-arrestin family members in C. elegans and their sequence similarity1, 7, which may obscure examining the functional importance of each α-arrestin. In addition, some α-arrestins may mostly function by modulating their interacting proteins6. In this case, genetic inhibition of α-arrestins may rather subtly affect the functions of their interacting partners that directly regulate physiology, such as aging and longevity, causing weak or no phenotypes. Thus, it will be interesting to test the effects of simultaneous inhibition of α-arrestins possibly through targeting the arrestin domain, and to identify and to functionally characterize proteins that bind C. elegans α-arrestins.”
The authors knockdown of 24 of the 29 alpha-arrestin genes. I wonder why they didn’t generate RNAi clones for the remaining 5, especially since they did generate 3 other constructs for their analysis. Potentially, the important alpha-arrestin genes is amongst those not examined.
> We appreciate the reviewers’ comment. We aimed at obtaining all the α arrestin RNAi clones that are not commercially available, except two predicted pseudogenes, arrd-20 and arrd-21 (Wormbase, version WS259). We successfully constructed three RNAi clones targeting arrd-11, arrd-17 and arrd-26, but could not obtain the other three RNAi clones, targeting arrd-12, arrd-22 and arrd-27. We addressed this issue in Methods for RNAi clones and in Discussion as follows,
In Methods, RNAi clones: “RNAi clones for arrd-12, arrd-22, and arrd-27 were not generated because of unknown technical problems and were not included in the lifespan screen. arrd-20 and arrd-21 are predicted to be pseudogenes (Wormbase, version WS259) and were excluded from our screen.”
In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
The authors did not report any other behavioural or morphological phenotypes observed by alpha-arrestin gene knockdown. Any information on observed phenotypes would be of interest to the research community.
> We thank the reviewer for the valuable comment. One noticeable phenotype we observed was that the eggs laid by worms fed with ttm-2 RNAi bacteria hatched more frequently than control eggs on FUdR-treated plates, although we did not quantify the results. We described this issue in Methods as follows,
In Methods, RNAi screen using lifespan assays: “Eggs laid by ttm-2 RNAi-treated worms hatched more frequently than control eggs on FUdR-containing plates in two independent lifespan experiments.”
Competing Interests:No competing interests were disclosed.Close
Author Response
(F1000Research Advisory Board Member)
14 Nov 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
14 Nov 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Roger Pocock,
We sincerely appreciate your valuable comments on our manuscript. We agree with all your points and revised our manuscript to describe the limitations of this study.
...
Continue readingDear Dr. Roger Pocock,
We sincerely appreciate your valuable comments on our manuscript. We agree with all your points and revised our manuscript to describe the limitations of this study. We would like to mention that the main motivation for publishing this manuscript is to provide our negative results to researchers who are interested in characterizing α-arrestins in C. elegans. Our point-by-point responses are described below. We hope you find our revised manuscript is suitable for publication in F1000Research.
In this manuscript Park et al. performed RNAi knockdown of 24 of the 29 alpha-arrestin genes in C. elegans to ask whether they are important for lifespan regulation in wild type and daf-2 mutant animals. Their data shows that, in their experimental conditions, most alpha-arrestin genes are individually not important for regulating lifespan.
Concerns:
One major issue with the study is whether RNAi knockdown of the alpha-arrestin genes is working. As some alpha-arrestins are expressed in the nervous system, which is refractory to RNAi, this is a concern. To determine whether the RNAi knockdown is working then Q-RTPCR should be performed. However, this is not a small task for 24 genes, therefore, further experiments using sensitized genetic backgrounds for RNAi or elevated temperatures may tease out phenotypes for some alpha-arrestin genes. This could particularly be performed for the alpha-arrestin genes that when knocked down engender a slight effect on lifespan.
> We agree with the reviewer’s comment that the RNAi knockdown efficiency is a major concern in interpreting our screen results. It would have been great if we had performed qRT-PCR analysis. However, as the reviewers pointed out, this is not a small task. We currently do not have any plan for following up this project and want to inform other researchers of our screen data. Therefore, we further emphasized the pitfall of our results in the Discussion as follows,
In Discussion: “The lifespan-regulatory functions of α-arrestins remain largely unexplored at the organism level. Here we showed that RNAi knockdown of individual C. elegans α-arrestins had small or no effects on lifespan in wild-type or daf-2 mutants. Our study has limitations that need to be considered for interpretation. First, RNAi targeting some α-arrestins might be insufficient for causing strong lifespan phenotypes. This may be because many of C. elegans α-arrestins are predicted to be expressed in neurons7, 9, 24-26, which are refractory to RNAi27-29. Lifespan assays using RNAi-hypersensitive mutants, including rrf-3(-) and eri-1(-) animals29-31, treated with α-arrestin RNAi, or using α-arrestin null mutants will help address this issue. In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR, our negative data should be interpreted with caution.”
In parallel to this, the authors could align the alpha-arrestin gene sequences to ask if there is a common sequence that could be used by RNAi targeting to knockdown multiple alpha-arrestin genes simultaneously.
> We appreciate the reviewer’s comment. The α-arrestin family members share arrestin-domains (O. Hobert, Wormbook, 2013). Thus, it will be intriguing if we could examine whether targeting arrestin domain for RNAi to knock down multiple arrd genes simultaneously and if multiple arrd gene knock down has effects on lifespan. We modified our manuscript to address this issue in Discussion as follows,
In Discussion: “Second, C. elegans α-arrestins may have functional redundancy considering the large number of the α-arrestin family members in C. elegans and their sequence similarity1, 7, which may obscure examining the functional importance of each α-arrestin. In addition, some α-arrestins may mostly function by modulating their interacting proteins6. In this case, genetic inhibition of α-arrestins may rather subtly affect the functions of their interacting partners that directly regulate physiology, such as aging and longevity, causing weak or no phenotypes. Thus, it will be interesting to test the effects of simultaneous inhibition of α-arrestins possibly through targeting the arrestin domain, and to identify and to functionally characterize proteins that bind C. elegans α-arrestins.”
The authors knockdown of 24 of the 29 alpha-arrestin genes. I wonder why they didn’t generate RNAi clones for the remaining 5, especially since they did generate 3 other constructs for their analysis. Potentially, the important alpha-arrestin genes is amongst those not examined.
> We appreciate the reviewers’ comment. We aimed at obtaining all the α arrestin RNAi clones that are not commercially available, except two predicted pseudogenes, arrd-20 and arrd-21 (Wormbase, version WS259). We successfully constructed three RNAi clones targeting arrd-11, arrd-17 and arrd-26, but could not obtain the other three RNAi clones, targeting arrd-12, arrd-22 and arrd-27. We addressed this issue in Methods for RNAi clones and in Discussion as follows,
In Methods, RNAi clones: “RNAi clones for arrd-12, arrd-22, and arrd-27 were not generated because of unknown technical problems and were not included in the lifespan screen. arrd-20 and arrd-21 are predicted to be pseudogenes (Wormbase, version WS259) and were excluded from our screen.”
In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
The authors did not report any other behavioural or morphological phenotypes observed by alpha-arrestin gene knockdown. Any information on observed phenotypes would be of interest to the research community.
> We thank the reviewer for the valuable comment. One noticeable phenotype we observed was that the eggs laid by worms fed with ttm-2 RNAi bacteria hatched more frequently than control eggs on FUdR-treated plates, although we did not quantify the results. We described this issue in Methods as follows,
In Methods, RNAi screen using lifespan assays: “Eggs laid by ttm-2 RNAi-treated worms hatched more frequently than control eggs on FUdR-containing plates in two independent lifespan experiments.”
Dear Dr. Roger Pocock,
We sincerely appreciate your valuable comments on our manuscript. We agree with all your points and revised our manuscript to describe the limitations of this study. We would like to mention that the main motivation for publishing this manuscript is to provide our negative results to researchers who are interested in characterizing α-arrestins in C. elegans. Our point-by-point responses are described below. We hope you find our revised manuscript is suitable for publication in F1000Research.
In this manuscript Park et al. performed RNAi knockdown of 24 of the 29 alpha-arrestin genes in C. elegans to ask whether they are important for lifespan regulation in wild type and daf-2 mutant animals. Their data shows that, in their experimental conditions, most alpha-arrestin genes are individually not important for regulating lifespan.
Concerns:
One major issue with the study is whether RNAi knockdown of the alpha-arrestin genes is working. As some alpha-arrestins are expressed in the nervous system, which is refractory to RNAi, this is a concern. To determine whether the RNAi knockdown is working then Q-RTPCR should be performed. However, this is not a small task for 24 genes, therefore, further experiments using sensitized genetic backgrounds for RNAi or elevated temperatures may tease out phenotypes for some alpha-arrestin genes. This could particularly be performed for the alpha-arrestin genes that when knocked down engender a slight effect on lifespan.
> We agree with the reviewer’s comment that the RNAi knockdown efficiency is a major concern in interpreting our screen results. It would have been great if we had performed qRT-PCR analysis. However, as the reviewers pointed out, this is not a small task. We currently do not have any plan for following up this project and want to inform other researchers of our screen data. Therefore, we further emphasized the pitfall of our results in the Discussion as follows,
In Discussion: “The lifespan-regulatory functions of α-arrestins remain largely unexplored at the organism level. Here we showed that RNAi knockdown of individual C. elegans α-arrestins had small or no effects on lifespan in wild-type or daf-2 mutants. Our study has limitations that need to be considered for interpretation. First, RNAi targeting some α-arrestins might be insufficient for causing strong lifespan phenotypes. This may be because many of C. elegans α-arrestins are predicted to be expressed in neurons7, 9, 24-26, which are refractory to RNAi27-29. Lifespan assays using RNAi-hypersensitive mutants, including rrf-3(-) and eri-1(-) animals29-31, treated with α-arrestin RNAi, or using α-arrestin null mutants will help address this issue. In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR, our negative data should be interpreted with caution.”
In parallel to this, the authors could align the alpha-arrestin gene sequences to ask if there is a common sequence that could be used by RNAi targeting to knockdown multiple alpha-arrestin genes simultaneously.
> We appreciate the reviewer’s comment. The α-arrestin family members share arrestin-domains (O. Hobert, Wormbook, 2013). Thus, it will be intriguing if we could examine whether targeting arrestin domain for RNAi to knock down multiple arrd genes simultaneously and if multiple arrd gene knock down has effects on lifespan. We modified our manuscript to address this issue in Discussion as follows,
In Discussion: “Second, C. elegans α-arrestins may have functional redundancy considering the large number of the α-arrestin family members in C. elegans and their sequence similarity1, 7, which may obscure examining the functional importance of each α-arrestin. In addition, some α-arrestins may mostly function by modulating their interacting proteins6. In this case, genetic inhibition of α-arrestins may rather subtly affect the functions of their interacting partners that directly regulate physiology, such as aging and longevity, causing weak or no phenotypes. Thus, it will be interesting to test the effects of simultaneous inhibition of α-arrestins possibly through targeting the arrestin domain, and to identify and to functionally characterize proteins that bind C. elegans α-arrestins.”
The authors knockdown of 24 of the 29 alpha-arrestin genes. I wonder why they didn’t generate RNAi clones for the remaining 5, especially since they did generate 3 other constructs for their analysis. Potentially, the important alpha-arrestin genes is amongst those not examined.
> We appreciate the reviewers’ comment. We aimed at obtaining all the α arrestin RNAi clones that are not commercially available, except two predicted pseudogenes, arrd-20 and arrd-21 (Wormbase, version WS259). We successfully constructed three RNAi clones targeting arrd-11, arrd-17 and arrd-26, but could not obtain the other three RNAi clones, targeting arrd-12, arrd-22 and arrd-27. We addressed this issue in Methods for RNAi clones and in Discussion as follows,
In Methods, RNAi clones: “RNAi clones for arrd-12, arrd-22, and arrd-27 were not generated because of unknown technical problems and were not included in the lifespan screen. arrd-20 and arrd-21 are predicted to be pseudogenes (Wormbase, version WS259) and were excluded from our screen.”
In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
The authors did not report any other behavioural or morphological phenotypes observed by alpha-arrestin gene knockdown. Any information on observed phenotypes would be of interest to the research community.
> We thank the reviewer for the valuable comment. One noticeable phenotype we observed was that the eggs laid by worms fed with ttm-2 RNAi bacteria hatched more frequently than control eggs on FUdR-treated plates, although we did not quantify the results. We described this issue in Methods as follows,
In Methods, RNAi screen using lifespan assays: “Eggs laid by ttm-2 RNAi-treated worms hatched more frequently than control eggs on FUdR-containing plates in two independent lifespan experiments.”
Competing Interests:No competing interests were disclosed.Close
Bitto A and Pitt J. Reviewer Report For: RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan [version 4; peer review: 3 approved]. F1000Research 2017, 6:1515 (https://doi.org/10.5256/f1000research.13357.r25290)
In the manuscript RNAi targeting Caenorhabditis elegans alpha-arrestins marginally affects lifespan Park and co-workers provide largely negative results from lifespan experiments following RNAi knockdown of most of the worm alpha-arrestins. While we welcome the effort to publish negative results, there is
... Continue reading
In the manuscript RNAi targeting Caenorhabditis elegans alpha-arrestins marginally affects lifespan Park and co-workers provide largely negative results from lifespan experiments following RNAi knockdown of most of the worm alpha-arrestins. While we welcome the effort to publish negative results, there is one major concern with the experimental approach and several minor concerns with the interpretation of the results that should be addressed before the manuscript is fully accepted for indexing in F1000Research.
Major concern:
The authors data is almost entirely negative: there were only 4 clones which had a statistically significant effect. The authors attempt to address this concern by providing RNAi data for daf-16, which is known to strongly suppress the lifespan extension of daf-2(e1370) as well as decreasing N2 lifespan. However, the authors must demonstrate that alpha-arrestin RNAi knockdown is functioning. As suggested by the authors, there is some evidence that alpha-arrestins are expressed in neurons which are notoriously refractory to RNAi. The authors must show that they can use RNAi to knockdown alpha-arrestins by QRTPCR analysis. The authors could also address this concern by performing the RNAi in one of the mutant strains in which RNAi works in neurons and report if the lifespan still does not change, as they suggest in the discussion. We believe that providing these data is necessary to correctly interpret the results from the lifespan analyses.
Minor concerns:
1. In the text the authors speak broadly of individual arrestins having no role in longevity however the authors only tested 24 of the 29 arrestins in the worm. If the authors had limited themselves to only those clones available in commercial RNAi libraries this would be understandable, however the authors did construct 3 new RNAi feeding constructs. This brings up the question to why the authors did not prepare the remaining 5 constructs. If the authors have reason to exclude those genes from their analysis (ie. they are pseudogenes, not expressed, etc) they should discuss these reasons in the text. Besides, there is the possibility that one of the missing constructs may indeed have major effects on lifespan in C. elegans.
2. The title of the manuscript implies there are more than one result than was observed. The simpler conclusion from the results provided is that individual alpha-arrestin knockdown has no effect of lifespan. This is the fundamental observation of the paper considering that most of them (20 out of 24) had no effect and that 4 of them had such an extremely small effect that it is arguable that the sample size is insufficient to support any conclusion1. We understand the authors' motivation to make the manuscript sound more appealing, however, their interpretation is disingenuous, given their results.
3. Because alpha-arrestins share significant sequence homology the authors should make some mention as to the likelihood that RNAi of any one arrestin would have off target effects on other arrestins. The authors could easily address this question experimentally with QRTPCR analysis. This is particularly relevant as one of the main conclusions of the manuscript is that future alpha-arrestin studies should knockdown multiple alpha-arrestins simultaneously.
4. Given that the authors observed the RNAi treated animals for weeks or months (in daf-2) it is surprising that no other behavioral or physical phenotypes from the alpha-arrestin RNAi were observed during the course of the analysis. If the authors truly did not observe any other phenotypes they should state it briefly in the text.
Is the work clearly and accurately presented and does it cite the current literature?
Partly
Is the study design appropriate and is the work technically sound?
Partly
Are sufficient details of methods and analysis provided to allow replication by others?
Yes
If applicable, is the statistical analysis and its interpretation appropriate?
Partly
Are all the source data underlying the results available to ensure full reproducibility?
Yes
Are the conclusions drawn adequately supported by the results?
No
References
1. Petrascheck M, Miller DL: Computational Analysis of Lifespan Experiment Reproducibility.Front Genet. 2017; 8: 92 PubMed Abstract | Publisher Full Text
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Biology of Aging
We confirm that we have read this submission and believe that we have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however we have significant reservations, as outlined above.
Bitto A and Pitt J. Reviewer Report For: RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan [version 4; peer review: 3 approved]. F1000Research 2017, 6:1515 (https://doi.org/10.5256/f1000research.13357.r25290)
Author Response
(F1000Research Advisory Board Member)
14 Nov 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
14 Nov 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Alessandro Bitto and Dr. Jason Pitt,
We greatly appreciate the reviewers’ valuable comments on our manuscript. We agree with all the reviewers’ comments and modified our manuscript to
...
Continue readingDear Dr. Alessandro Bitto and Dr. Jason Pitt,
We greatly appreciate the reviewers’ valuable comments on our manuscript. We agree with all the reviewers’ comments and modified our manuscript to describe the limitations of our studies raised by the reviewers. We would like to mention that the main purpose of publishing our current manuscript is to provide our data to other researchers who are interested in the characterization of C. elegansarrd genes, instead of keeping our current data unpublished. Our point-by-point responses are described below. We hope you find our revised manuscript is suitable for publication in F1000Research.
Reviewers’ comments:
In the manuscript RNAi targeting Caenorhabditis elegans alpha-arrestins marginally affects lifespan Park and co-workers provide largely negative results from lifespan experiments following RNAi knockdown of most of the worm alpha-arrestins. While we welcome the effort to publish negative results, there is one major concern with the experimental approach and several minor concerns with the interpretation of the results that should be addressed before the manuscript is fully accepted for indexing in F1000Research.
Major concern:
The authors data is almost entirely negative: there were only 4 clones which had a statistically significant effect. The authors attempt to address this concern by providing RNAi data for daf-16, which is known to strongly suppress the lifespan extension of daf-2(e1370) as well as decreasing N2 lifespan. However, the authors must demonstrate that alpha-arrestin RNAi knockdown is functioning. As suggested by the authors, there is some evidence that alpha-arrestins are expressed in neurons which are notoriously refractory to RNAi. The authors must show that they can use RNAi to knockdown alpha-arrestins by QRTPCR analysis. The authors could also address this concern by performing the RNAi in one of the mutant strains in which RNAi works in neurons and report if the lifespan still does not change, as they suggest in the discussion. We believe that providing these data is necessary to correctly interpret the results from the lifespan analyses.
> We completely agree with the reviewers for the comment. It would have been great if we had performed qRTPCR for validating the RNAi efficiency, while we were doing lifespan assays. As our experiments were done as an RNAi screen, our initial plan was validating RNAi efficiency and using RNAi-sensitive mutants if we had identified some positive RNAi clones. However, as the reviewers pointed out, our screen results were negative. We currently do not have any plan for following up this project and just want to inform other researchers of our screen data. Therefore, instead of putting efforts on this project retrospectively, we emphasized the limitations of our data raised by the reviewers to inform the readers of pitfalls by revising the manuscript as follows,
In Discussion: “The lifespan-regulatory functions of α-arrestins remain largely unexplored at the organism level. Here we showed that RNAi knockdown of individual C. elegans α-arrestins had small or no effects on lifespan in wild-type or daf-2 mutants. Our study has limitations that need to be considered for interpretation. First, RNAi targeting some α-arrestins might be insufficient for causing strong lifespan phenotypes. This may be because many of C. elegans α-arrestins are predicted to be expressed in neurons7, 9, 24-26, which are refractory to RNAi 27-29. Lifespan assays using RNAi-hypersensitive mutants, including rrf-3(-) and eri-1(-) animals 29-31, treated with α-arrestin RNAi, or using α-arrestin null mutants will help address this issue. In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR, our negative data should be interpreted with caution.”
Minor concerns:
1. In the text the authors speak broadly of individual arrestins having no role in longevity however the authors only tested 24 of the 29 arrestins in the worm. If the authors had limited themselves to only those clones available in commercial RNAi libraries this would be understandable, however the authors did construct 3 new RNAi feeding constructs. This brings up the question to why the authors did not prepare the remaining 5 constructs. If the authors have reason to exclude those genes from their analysis (ie. they are pseudogenes, not expressed, etc) they should discuss these reasons in the text. Besides, there is the possibility that one of the missing constructs may indeed have major effects on lifespan in C. elegans.
> We appreciate the reviewers’ comment. We initially aimed at obtaining all the α arrestin RNAi clones that are not commercially available except two predicted pseudogenes, arrd-20 and arrd-21 (Wormbase, version WS259). We successfully constructed three RNAi clones targeting arrd-11, arrd-17 and arrd-26, but could not obtain the other three RNAi clones for arrd-12, arrd-22 and arrd-27 because of unknown technical problems. We agree with the reviewers’ point that these three genes that were not tested may have major effects on lifespan. We addressed this issue in Methods for RNAi clones as well as in Discussion as follows,
In Methods, RNAi clones: “RNAi clones for arrd-12, arrd-22, and arrd-27 were not generated because of unknown technical problems and were not included in the lifespan screen. arrd-20 and arrd-21 are predicted to be pseudogenes (Wormbase, version WS259) and were excluded from our screen.”
In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
2. The title of the manuscript implies there are more than one result than was observed. The simpler conclusion from the results provided is that individual alpha-arrestin knockdown has no effect of lifespan. This is the fundamental observation of the paper considering that most of them (20 out of 24) had no effect and that 4 of them had such an extremely small effect that it is arguable that the sample size is insufficient to support any conclusion1. We understand the authors' motivation to make the manuscript sound more appealing, however, their interpretation is disingenuous, given their results.
> We thank the reviewers’ comment on the title and changed the title as follows,
Title: “RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan”
3. Because alpha-arrestins share significant sequence homology the authors should make some mention as to the likelihood that RNAi of any one arrestin would have off target effects on other arrestins. The authors could easily address this question experimentally with QRTPCR analysis. This is particularly relevant as one of the main conclusions of the manuscript is that future alpha-arrestin studies should knockdown multiple alpha-arrestins simultaneously.
> We agree with the reviewers that the RNAi targeting each of α-arrestin genes may have caused off-target effects on other α-arrestin genes because of sequence similarities. We performed off-target prediction using Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap), and at least with the prediction, the RNAi clones used in our screen do not appear to have major off-target effects on any other genes including α-arrestin genes, except arrd-19 RNAi which may also target arrd-21 which is a predicted pseudogene. Although qRT-PCR will be a straightforward way as the reviewers suggested, again currently we do not have any plan on following up this project and we therefore instead acknowledged the limitation. We added the prediction results in the Supplementary Table S1 and described this issue in the Methods, RNAi clones part as follows,
In Methods, RNAi clones: “All RNAi clones used in our screen were examined for their potential off-target effects by using Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap)22, and no significant off-target was predicted except arrd-19 RNAi, which may additionally target a predicted pseudogene arrd-21 (Table S1). Experimental validation by qRT-PCR will be necessary to completely exclude the possible off-target effects by RNAi clones.”
In Supplementary material: “Table S1. Possible RNAi target prediction. RNAi clones used in the screen were examined for their possible off-targets by Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap)22. For RNAi clone names, ‘mv’ indicates a RNAi clone from Vidal library and ‘sjj’ indicates a RNAi clone from Ahringer library. RNAi clones generated by us for this study are labeled as ‘Lee lab’. Scores for potential target transcripts were calculated as described previously22 and were rounded off to the second digit after the decimal point.”
In Reference: 22. Thakur, N., et al., Clone Mapper: An Online Suite of Tools for RNAi Experiments in Caenorhabditis elegans. G3 (Bethesda), 2014. 4(11): p. 2137-2145.23.
4. Given that the authors observed the RNAi treated animals for weeks or months (in daf-2) it is surprising that no other behavioral or physical phenotypes from the alpha-arrestin RNAi were observed during the course of the analysis. If the authors truly did not observe any other phenotypes they should state it briefly in the text.
> We appreciate the reviewers’ comment. As our main interests were lifespan changes from a large scale screen, we did not pay attention to other phenotypes. Nevertheless, one noticeable phenotype we observed was that the eggs laid by worms fed with ttm-2 RNAi bacteria hatched more frequently than control eggs on FUdR-treated plates, although we did not quantitate the results. We described this issue in Methods as follows,
In Methods, RNAi screen using lifespan assays: “Eggs laid by ttm-2 RNAi-treated worms hatched more frequently than control eggs on FUdR-containing plates in two independent lifespan experiments.”
Dear Dr. Alessandro Bitto and Dr. Jason Pitt,
We greatly appreciate the reviewers’ valuable comments on our manuscript. We agree with all the reviewers’ comments and modified our manuscript to describe the limitations of our studies raised by the reviewers. We would like to mention that the main purpose of publishing our current manuscript is to provide our data to other researchers who are interested in the characterization of C. elegansarrd genes, instead of keeping our current data unpublished. Our point-by-point responses are described below. We hope you find our revised manuscript is suitable for publication in F1000Research.
Reviewers’ comments:
In the manuscript RNAi targeting Caenorhabditis elegans alpha-arrestins marginally affects lifespan Park and co-workers provide largely negative results from lifespan experiments following RNAi knockdown of most of the worm alpha-arrestins. While we welcome the effort to publish negative results, there is one major concern with the experimental approach and several minor concerns with the interpretation of the results that should be addressed before the manuscript is fully accepted for indexing in F1000Research.
Major concern:
The authors data is almost entirely negative: there were only 4 clones which had a statistically significant effect. The authors attempt to address this concern by providing RNAi data for daf-16, which is known to strongly suppress the lifespan extension of daf-2(e1370) as well as decreasing N2 lifespan. However, the authors must demonstrate that alpha-arrestin RNAi knockdown is functioning. As suggested by the authors, there is some evidence that alpha-arrestins are expressed in neurons which are notoriously refractory to RNAi. The authors must show that they can use RNAi to knockdown alpha-arrestins by QRTPCR analysis. The authors could also address this concern by performing the RNAi in one of the mutant strains in which RNAi works in neurons and report if the lifespan still does not change, as they suggest in the discussion. We believe that providing these data is necessary to correctly interpret the results from the lifespan analyses.
> We completely agree with the reviewers for the comment. It would have been great if we had performed qRTPCR for validating the RNAi efficiency, while we were doing lifespan assays. As our experiments were done as an RNAi screen, our initial plan was validating RNAi efficiency and using RNAi-sensitive mutants if we had identified some positive RNAi clones. However, as the reviewers pointed out, our screen results were negative. We currently do not have any plan for following up this project and just want to inform other researchers of our screen data. Therefore, instead of putting efforts on this project retrospectively, we emphasized the limitations of our data raised by the reviewers to inform the readers of pitfalls by revising the manuscript as follows,
In Discussion: “The lifespan-regulatory functions of α-arrestins remain largely unexplored at the organism level. Here we showed that RNAi knockdown of individual C. elegans α-arrestins had small or no effects on lifespan in wild-type or daf-2 mutants. Our study has limitations that need to be considered for interpretation. First, RNAi targeting some α-arrestins might be insufficient for causing strong lifespan phenotypes. This may be because many of C. elegans α-arrestins are predicted to be expressed in neurons7, 9, 24-26, which are refractory to RNAi 27-29. Lifespan assays using RNAi-hypersensitive mutants, including rrf-3(-) and eri-1(-) animals 29-31, treated with α-arrestin RNAi, or using α-arrestin null mutants will help address this issue. In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR, our negative data should be interpreted with caution.”
Minor concerns:
1. In the text the authors speak broadly of individual arrestins having no role in longevity however the authors only tested 24 of the 29 arrestins in the worm. If the authors had limited themselves to only those clones available in commercial RNAi libraries this would be understandable, however the authors did construct 3 new RNAi feeding constructs. This brings up the question to why the authors did not prepare the remaining 5 constructs. If the authors have reason to exclude those genes from their analysis (ie. they are pseudogenes, not expressed, etc) they should discuss these reasons in the text. Besides, there is the possibility that one of the missing constructs may indeed have major effects on lifespan in C. elegans.
> We appreciate the reviewers’ comment. We initially aimed at obtaining all the α arrestin RNAi clones that are not commercially available except two predicted pseudogenes, arrd-20 and arrd-21 (Wormbase, version WS259). We successfully constructed three RNAi clones targeting arrd-11, arrd-17 and arrd-26, but could not obtain the other three RNAi clones for arrd-12, arrd-22 and arrd-27 because of unknown technical problems. We agree with the reviewers’ point that these three genes that were not tested may have major effects on lifespan. We addressed this issue in Methods for RNAi clones as well as in Discussion as follows,
In Methods, RNAi clones: “RNAi clones for arrd-12, arrd-22, and arrd-27 were not generated because of unknown technical problems and were not included in the lifespan screen. arrd-20 and arrd-21 are predicted to be pseudogenes (Wormbase, version WS259) and were excluded from our screen.”
In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
2. The title of the manuscript implies there are more than one result than was observed. The simpler conclusion from the results provided is that individual alpha-arrestin knockdown has no effect of lifespan. This is the fundamental observation of the paper considering that most of them (20 out of 24) had no effect and that 4 of them had such an extremely small effect that it is arguable that the sample size is insufficient to support any conclusion1. We understand the authors' motivation to make the manuscript sound more appealing, however, their interpretation is disingenuous, given their results.
> We thank the reviewers’ comment on the title and changed the title as follows,
Title: “RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan”
3. Because alpha-arrestins share significant sequence homology the authors should make some mention as to the likelihood that RNAi of any one arrestin would have off target effects on other arrestins. The authors could easily address this question experimentally with QRTPCR analysis. This is particularly relevant as one of the main conclusions of the manuscript is that future alpha-arrestin studies should knockdown multiple alpha-arrestins simultaneously.
> We agree with the reviewers that the RNAi targeting each of α-arrestin genes may have caused off-target effects on other α-arrestin genes because of sequence similarities. We performed off-target prediction using Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap), and at least with the prediction, the RNAi clones used in our screen do not appear to have major off-target effects on any other genes including α-arrestin genes, except arrd-19 RNAi which may also target arrd-21 which is a predicted pseudogene. Although qRT-PCR will be a straightforward way as the reviewers suggested, again currently we do not have any plan on following up this project and we therefore instead acknowledged the limitation. We added the prediction results in the Supplementary Table S1 and described this issue in the Methods, RNAi clones part as follows,
In Methods, RNAi clones: “All RNAi clones used in our screen were examined for their potential off-target effects by using Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap)22, and no significant off-target was predicted except arrd-19 RNAi, which may additionally target a predicted pseudogene arrd-21 (Table S1). Experimental validation by qRT-PCR will be necessary to completely exclude the possible off-target effects by RNAi clones.”
In Supplementary material: “Table S1. Possible RNAi target prediction. RNAi clones used in the screen were examined for their possible off-targets by Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap)22. For RNAi clone names, ‘mv’ indicates a RNAi clone from Vidal library and ‘sjj’ indicates a RNAi clone from Ahringer library. RNAi clones generated by us for this study are labeled as ‘Lee lab’. Scores for potential target transcripts were calculated as described previously22 and were rounded off to the second digit after the decimal point.”
In Reference: 22. Thakur, N., et al., Clone Mapper: An Online Suite of Tools for RNAi Experiments in Caenorhabditis elegans. G3 (Bethesda), 2014. 4(11): p. 2137-2145.23.
4. Given that the authors observed the RNAi treated animals for weeks or months (in daf-2) it is surprising that no other behavioral or physical phenotypes from the alpha-arrestin RNAi were observed during the course of the analysis. If the authors truly did not observe any other phenotypes they should state it briefly in the text.
> We appreciate the reviewers’ comment. As our main interests were lifespan changes from a large scale screen, we did not pay attention to other phenotypes. Nevertheless, one noticeable phenotype we observed was that the eggs laid by worms fed with ttm-2 RNAi bacteria hatched more frequently than control eggs on FUdR-treated plates, although we did not quantitate the results. We described this issue in Methods as follows,
In Methods, RNAi screen using lifespan assays: “Eggs laid by ttm-2 RNAi-treated worms hatched more frequently than control eggs on FUdR-containing plates in two independent lifespan experiments.”
Competing Interests:No competing interests were disclosed.Close
Author Response
(F1000Research Advisory Board Member)
14 Nov 2017
Seung-Jae V Lee, Korea Advanced Institute of Science and Technology, South Korea
14 Nov 2017
Author Response F1000Research Advisory Board Member
Dear Dr. Alessandro Bitto and Dr. Jason Pitt,
We greatly appreciate the reviewers’ valuable comments on our manuscript. We agree with all the reviewers’ comments and modified our manuscript to
...
Continue readingDear Dr. Alessandro Bitto and Dr. Jason Pitt,
We greatly appreciate the reviewers’ valuable comments on our manuscript. We agree with all the reviewers’ comments and modified our manuscript to describe the limitations of our studies raised by the reviewers. We would like to mention that the main purpose of publishing our current manuscript is to provide our data to other researchers who are interested in the characterization of C. elegansarrd genes, instead of keeping our current data unpublished. Our point-by-point responses are described below. We hope you find our revised manuscript is suitable for publication in F1000Research.
Reviewers’ comments:
In the manuscript RNAi targeting Caenorhabditis elegans alpha-arrestins marginally affects lifespan Park and co-workers provide largely negative results from lifespan experiments following RNAi knockdown of most of the worm alpha-arrestins. While we welcome the effort to publish negative results, there is one major concern with the experimental approach and several minor concerns with the interpretation of the results that should be addressed before the manuscript is fully accepted for indexing in F1000Research.
Major concern:
The authors data is almost entirely negative: there were only 4 clones which had a statistically significant effect. The authors attempt to address this concern by providing RNAi data for daf-16, which is known to strongly suppress the lifespan extension of daf-2(e1370) as well as decreasing N2 lifespan. However, the authors must demonstrate that alpha-arrestin RNAi knockdown is functioning. As suggested by the authors, there is some evidence that alpha-arrestins are expressed in neurons which are notoriously refractory to RNAi. The authors must show that they can use RNAi to knockdown alpha-arrestins by QRTPCR analysis. The authors could also address this concern by performing the RNAi in one of the mutant strains in which RNAi works in neurons and report if the lifespan still does not change, as they suggest in the discussion. We believe that providing these data is necessary to correctly interpret the results from the lifespan analyses.
> We completely agree with the reviewers for the comment. It would have been great if we had performed qRTPCR for validating the RNAi efficiency, while we were doing lifespan assays. As our experiments were done as an RNAi screen, our initial plan was validating RNAi efficiency and using RNAi-sensitive mutants if we had identified some positive RNAi clones. However, as the reviewers pointed out, our screen results were negative. We currently do not have any plan for following up this project and just want to inform other researchers of our screen data. Therefore, instead of putting efforts on this project retrospectively, we emphasized the limitations of our data raised by the reviewers to inform the readers of pitfalls by revising the manuscript as follows,
In Discussion: “The lifespan-regulatory functions of α-arrestins remain largely unexplored at the organism level. Here we showed that RNAi knockdown of individual C. elegans α-arrestins had small or no effects on lifespan in wild-type or daf-2 mutants. Our study has limitations that need to be considered for interpretation. First, RNAi targeting some α-arrestins might be insufficient for causing strong lifespan phenotypes. This may be because many of C. elegans α-arrestins are predicted to be expressed in neurons7, 9, 24-26, which are refractory to RNAi 27-29. Lifespan assays using RNAi-hypersensitive mutants, including rrf-3(-) and eri-1(-) animals 29-31, treated with α-arrestin RNAi, or using α-arrestin null mutants will help address this issue. In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR, our negative data should be interpreted with caution.”
Minor concerns:
1. In the text the authors speak broadly of individual arrestins having no role in longevity however the authors only tested 24 of the 29 arrestins in the worm. If the authors had limited themselves to only those clones available in commercial RNAi libraries this would be understandable, however the authors did construct 3 new RNAi feeding constructs. This brings up the question to why the authors did not prepare the remaining 5 constructs. If the authors have reason to exclude those genes from their analysis (ie. they are pseudogenes, not expressed, etc) they should discuss these reasons in the text. Besides, there is the possibility that one of the missing constructs may indeed have major effects on lifespan in C. elegans.
> We appreciate the reviewers’ comment. We initially aimed at obtaining all the α arrestin RNAi clones that are not commercially available except two predicted pseudogenes, arrd-20 and arrd-21 (Wormbase, version WS259). We successfully constructed three RNAi clones targeting arrd-11, arrd-17 and arrd-26, but could not obtain the other three RNAi clones for arrd-12, arrd-22 and arrd-27 because of unknown technical problems. We agree with the reviewers’ point that these three genes that were not tested may have major effects on lifespan. We addressed this issue in Methods for RNAi clones as well as in Discussion as follows,
In Methods, RNAi clones: “RNAi clones for arrd-12, arrd-22, and arrd-27 were not generated because of unknown technical problems and were not included in the lifespan screen. arrd-20 and arrd-21 are predicted to be pseudogenes (Wormbase, version WS259) and were excluded from our screen.”
In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
2. The title of the manuscript implies there are more than one result than was observed. The simpler conclusion from the results provided is that individual alpha-arrestin knockdown has no effect of lifespan. This is the fundamental observation of the paper considering that most of them (20 out of 24) had no effect and that 4 of them had such an extremely small effect that it is arguable that the sample size is insufficient to support any conclusion1. We understand the authors' motivation to make the manuscript sound more appealing, however, their interpretation is disingenuous, given their results.
> We thank the reviewers’ comment on the title and changed the title as follows,
Title: “RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan”
3. Because alpha-arrestins share significant sequence homology the authors should make some mention as to the likelihood that RNAi of any one arrestin would have off target effects on other arrestins. The authors could easily address this question experimentally with QRTPCR analysis. This is particularly relevant as one of the main conclusions of the manuscript is that future alpha-arrestin studies should knockdown multiple alpha-arrestins simultaneously.
> We agree with the reviewers that the RNAi targeting each of α-arrestin genes may have caused off-target effects on other α-arrestin genes because of sequence similarities. We performed off-target prediction using Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap), and at least with the prediction, the RNAi clones used in our screen do not appear to have major off-target effects on any other genes including α-arrestin genes, except arrd-19 RNAi which may also target arrd-21 which is a predicted pseudogene. Although qRT-PCR will be a straightforward way as the reviewers suggested, again currently we do not have any plan on following up this project and we therefore instead acknowledged the limitation. We added the prediction results in the Supplementary Table S1 and described this issue in the Methods, RNAi clones part as follows,
In Methods, RNAi clones: “All RNAi clones used in our screen were examined for their potential off-target effects by using Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap)22, and no significant off-target was predicted except arrd-19 RNAi, which may additionally target a predicted pseudogene arrd-21 (Table S1). Experimental validation by qRT-PCR will be necessary to completely exclude the possible off-target effects by RNAi clones.”
In Supplementary material: “Table S1. Possible RNAi target prediction. RNAi clones used in the screen were examined for their possible off-targets by Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap)22. For RNAi clone names, ‘mv’ indicates a RNAi clone from Vidal library and ‘sjj’ indicates a RNAi clone from Ahringer library. RNAi clones generated by us for this study are labeled as ‘Lee lab’. Scores for potential target transcripts were calculated as described previously22 and were rounded off to the second digit after the decimal point.”
In Reference: 22. Thakur, N., et al., Clone Mapper: An Online Suite of Tools for RNAi Experiments in Caenorhabditis elegans. G3 (Bethesda), 2014. 4(11): p. 2137-2145.23.
4. Given that the authors observed the RNAi treated animals for weeks or months (in daf-2) it is surprising that no other behavioral or physical phenotypes from the alpha-arrestin RNAi were observed during the course of the analysis. If the authors truly did not observe any other phenotypes they should state it briefly in the text.
> We appreciate the reviewers’ comment. As our main interests were lifespan changes from a large scale screen, we did not pay attention to other phenotypes. Nevertheless, one noticeable phenotype we observed was that the eggs laid by worms fed with ttm-2 RNAi bacteria hatched more frequently than control eggs on FUdR-treated plates, although we did not quantitate the results. We described this issue in Methods as follows,
In Methods, RNAi screen using lifespan assays: “Eggs laid by ttm-2 RNAi-treated worms hatched more frequently than control eggs on FUdR-containing plates in two independent lifespan experiments.”
Dear Dr. Alessandro Bitto and Dr. Jason Pitt,
We greatly appreciate the reviewers’ valuable comments on our manuscript. We agree with all the reviewers’ comments and modified our manuscript to describe the limitations of our studies raised by the reviewers. We would like to mention that the main purpose of publishing our current manuscript is to provide our data to other researchers who are interested in the characterization of C. elegansarrd genes, instead of keeping our current data unpublished. Our point-by-point responses are described below. We hope you find our revised manuscript is suitable for publication in F1000Research.
Reviewers’ comments:
In the manuscript RNAi targeting Caenorhabditis elegans alpha-arrestins marginally affects lifespan Park and co-workers provide largely negative results from lifespan experiments following RNAi knockdown of most of the worm alpha-arrestins. While we welcome the effort to publish negative results, there is one major concern with the experimental approach and several minor concerns with the interpretation of the results that should be addressed before the manuscript is fully accepted for indexing in F1000Research.
Major concern:
The authors data is almost entirely negative: there were only 4 clones which had a statistically significant effect. The authors attempt to address this concern by providing RNAi data for daf-16, which is known to strongly suppress the lifespan extension of daf-2(e1370) as well as decreasing N2 lifespan. However, the authors must demonstrate that alpha-arrestin RNAi knockdown is functioning. As suggested by the authors, there is some evidence that alpha-arrestins are expressed in neurons which are notoriously refractory to RNAi. The authors must show that they can use RNAi to knockdown alpha-arrestins by QRTPCR analysis. The authors could also address this concern by performing the RNAi in one of the mutant strains in which RNAi works in neurons and report if the lifespan still does not change, as they suggest in the discussion. We believe that providing these data is necessary to correctly interpret the results from the lifespan analyses.
> We completely agree with the reviewers for the comment. It would have been great if we had performed qRTPCR for validating the RNAi efficiency, while we were doing lifespan assays. As our experiments were done as an RNAi screen, our initial plan was validating RNAi efficiency and using RNAi-sensitive mutants if we had identified some positive RNAi clones. However, as the reviewers pointed out, our screen results were negative. We currently do not have any plan for following up this project and just want to inform other researchers of our screen data. Therefore, instead of putting efforts on this project retrospectively, we emphasized the limitations of our data raised by the reviewers to inform the readers of pitfalls by revising the manuscript as follows,
In Discussion: “The lifespan-regulatory functions of α-arrestins remain largely unexplored at the organism level. Here we showed that RNAi knockdown of individual C. elegans α-arrestins had small or no effects on lifespan in wild-type or daf-2 mutants. Our study has limitations that need to be considered for interpretation. First, RNAi targeting some α-arrestins might be insufficient for causing strong lifespan phenotypes. This may be because many of C. elegans α-arrestins are predicted to be expressed in neurons7, 9, 24-26, which are refractory to RNAi 27-29. Lifespan assays using RNAi-hypersensitive mutants, including rrf-3(-) and eri-1(-) animals 29-31, treated with α-arrestin RNAi, or using α-arrestin null mutants will help address this issue. In addition, as we did not test whether RNAi targeting each of α-arrestin genes was effective by using quantitative RT-PCR, our negative data should be interpreted with caution.”
Minor concerns:
1. In the text the authors speak broadly of individual arrestins having no role in longevity however the authors only tested 24 of the 29 arrestins in the worm. If the authors had limited themselves to only those clones available in commercial RNAi libraries this would be understandable, however the authors did construct 3 new RNAi feeding constructs. This brings up the question to why the authors did not prepare the remaining 5 constructs. If the authors have reason to exclude those genes from their analysis (ie. they are pseudogenes, not expressed, etc) they should discuss these reasons in the text. Besides, there is the possibility that one of the missing constructs may indeed have major effects on lifespan in C. elegans.
> We appreciate the reviewers’ comment. We initially aimed at obtaining all the α arrestin RNAi clones that are not commercially available except two predicted pseudogenes, arrd-20 and arrd-21 (Wormbase, version WS259). We successfully constructed three RNAi clones targeting arrd-11, arrd-17 and arrd-26, but could not obtain the other three RNAi clones for arrd-12, arrd-22 and arrd-27 because of unknown technical problems. We agree with the reviewers’ point that these three genes that were not tested may have major effects on lifespan. We addressed this issue in Methods for RNAi clones as well as in Discussion as follows,
In Methods, RNAi clones: “RNAi clones for arrd-12, arrd-22, and arrd-27 were not generated because of unknown technical problems and were not included in the lifespan screen. arrd-20 and arrd-21 are predicted to be pseudogenes (Wormbase, version WS259) and were excluded from our screen.”
In Discussion: “Third, it is possible that three α-arrestins, arrd-12, arrd-22 and arrd-27, which were not tested in our screen, may play crucial roles in lifespan regulation. Thus, it will be important to examine if genetic inhibition of each of these three α-arrestin genes affects lifespan in future studies.”
2. The title of the manuscript implies there are more than one result than was observed. The simpler conclusion from the results provided is that individual alpha-arrestin knockdown has no effect of lifespan. This is the fundamental observation of the paper considering that most of them (20 out of 24) had no effect and that 4 of them had such an extremely small effect that it is arguable that the sample size is insufficient to support any conclusion1. We understand the authors' motivation to make the manuscript sound more appealing, however, their interpretation is disingenuous, given their results.
> We thank the reviewers’ comment on the title and changed the title as follows,
Title: “RNAi targeting Caenorhabditis elegans α-arrestins has small or no effects on lifespan”
3. Because alpha-arrestins share significant sequence homology the authors should make some mention as to the likelihood that RNAi of any one arrestin would have off target effects on other arrestins. The authors could easily address this question experimentally with QRTPCR analysis. This is particularly relevant as one of the main conclusions of the manuscript is that future alpha-arrestin studies should knockdown multiple alpha-arrestins simultaneously.
> We agree with the reviewers that the RNAi targeting each of α-arrestin genes may have caused off-target effects on other α-arrestin genes because of sequence similarities. We performed off-target prediction using Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap), and at least with the prediction, the RNAi clones used in our screen do not appear to have major off-target effects on any other genes including α-arrestin genes, except arrd-19 RNAi which may also target arrd-21 which is a predicted pseudogene. Although qRT-PCR will be a straightforward way as the reviewers suggested, again currently we do not have any plan on following up this project and we therefore instead acknowledged the limitation. We added the prediction results in the Supplementary Table S1 and described this issue in the Methods, RNAi clones part as follows,
In Methods, RNAi clones: “All RNAi clones used in our screen were examined for their potential off-target effects by using Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap)22, and no significant off-target was predicted except arrd-19 RNAi, which may additionally target a predicted pseudogene arrd-21 (Table S1). Experimental validation by qRT-PCR will be necessary to completely exclude the possible off-target effects by RNAi clones.”
In Supplementary material: “Table S1. Possible RNAi target prediction. RNAi clones used in the screen were examined for their possible off-targets by Clone Mapper (http://bioinformatics.lif.univ-mrs.fr/RNAiMap)22. For RNAi clone names, ‘mv’ indicates a RNAi clone from Vidal library and ‘sjj’ indicates a RNAi clone from Ahringer library. RNAi clones generated by us for this study are labeled as ‘Lee lab’. Scores for potential target transcripts were calculated as described previously22 and were rounded off to the second digit after the decimal point.”
In Reference: 22. Thakur, N., et al., Clone Mapper: An Online Suite of Tools for RNAi Experiments in Caenorhabditis elegans. G3 (Bethesda), 2014. 4(11): p. 2137-2145.23.
4. Given that the authors observed the RNAi treated animals for weeks or months (in daf-2) it is surprising that no other behavioral or physical phenotypes from the alpha-arrestin RNAi were observed during the course of the analysis. If the authors truly did not observe any other phenotypes they should state it briefly in the text.
> We appreciate the reviewers’ comment. As our main interests were lifespan changes from a large scale screen, we did not pay attention to other phenotypes. Nevertheless, one noticeable phenotype we observed was that the eggs laid by worms fed with ttm-2 RNAi bacteria hatched more frequently than control eggs on FUdR-treated plates, although we did not quantitate the results. We described this issue in Methods as follows,
In Methods, RNAi screen using lifespan assays: “Eggs laid by ttm-2 RNAi-treated worms hatched more frequently than control eggs on FUdR-containing plates in two independent lifespan experiments.”
Competing Interests:No competing interests were disclosed.Close
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Spreadsheet data files may not format correctly if your computer is using different default delimiters (symbols used to separate values into separate cells) - a spreadsheet created in one region is sometimes misinterpreted by computers in other regions. You can change the regional settings on your computer so that the spreadsheet can be interpreted correctly.
How to fix it
Save downloaded CSV file
Open spreadsheet program (e.g. Excel)
Click the ‘Data’ tab at the top
Click the ‘From text’ icon (top left)
Browse for downloaded CSV file, click ‘Import’
Ensure ‘Delimited’ radio button is selected, click ‘Next’
Check one of the appropriate delimiter checkboxes (you can visualize the formatting by looking at the data preview below these options)
Park S, Jung Y, An SWA et al.. Dataset 1 in: RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan. F1000Research 2017, 6:1515 (https://doi.org/10.5256/f1000research.12337.d173158)
Spreadsheet data files may not format correctly if your computer is using different default delimiters (symbols used to separate values into separate cells) - a spreadsheet created in one region is sometimes misinterpreted by computers in other regions. You can change the regional settings on your computer so that the spreadsheet can be interpreted correctly.
How to fix it
Save downloaded CSV file
Open spreadsheet program (e.g. Excel)
Click the ‘Data’ tab at the top
Click the ‘From text’ icon (top left)
Browse for downloaded CSV file, click ‘Import’
Ensure ‘Delimited’ radio button is selected, click ‘Next’
Check one of the appropriate delimiter checkboxes (you can visualize the formatting by looking at the data preview below these options)
Park S, Jung Y, An SWA et al.. Dataset 2 in: RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan. F1000Research 2017, 6:1515 (https://doi.org/10.5256/f1000research.12337.d173159)
Spreadsheet data files may not format correctly if your computer is using different default delimiters (symbols used to separate values into separate cells) - a spreadsheet created in one region is sometimes misinterpreted by computers in other regions. You can change the regional settings on your computer so that the spreadsheet can be interpreted correctly.
How to fix it
Save downloaded CSV file
Open spreadsheet program (e.g. Excel)
Click the ‘Data’ tab at the top
Click the ‘From text’ icon (top left)
Browse for downloaded CSV file, click ‘Import’
Ensure ‘Delimited’ radio button is selected, click ‘Next’
Check one of the appropriate delimiter checkboxes (you can visualize the formatting by looking at the data preview below these options)
Park S, Jung Y, An SWA et al.. Dataset 3 in: RNAi targeting Caenorhabditis elegans α-arrestins has little effect on lifespan. F1000Research 2017, 6:1515 (https://doi.org/10.5256/f1000research.12337.d173160)
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
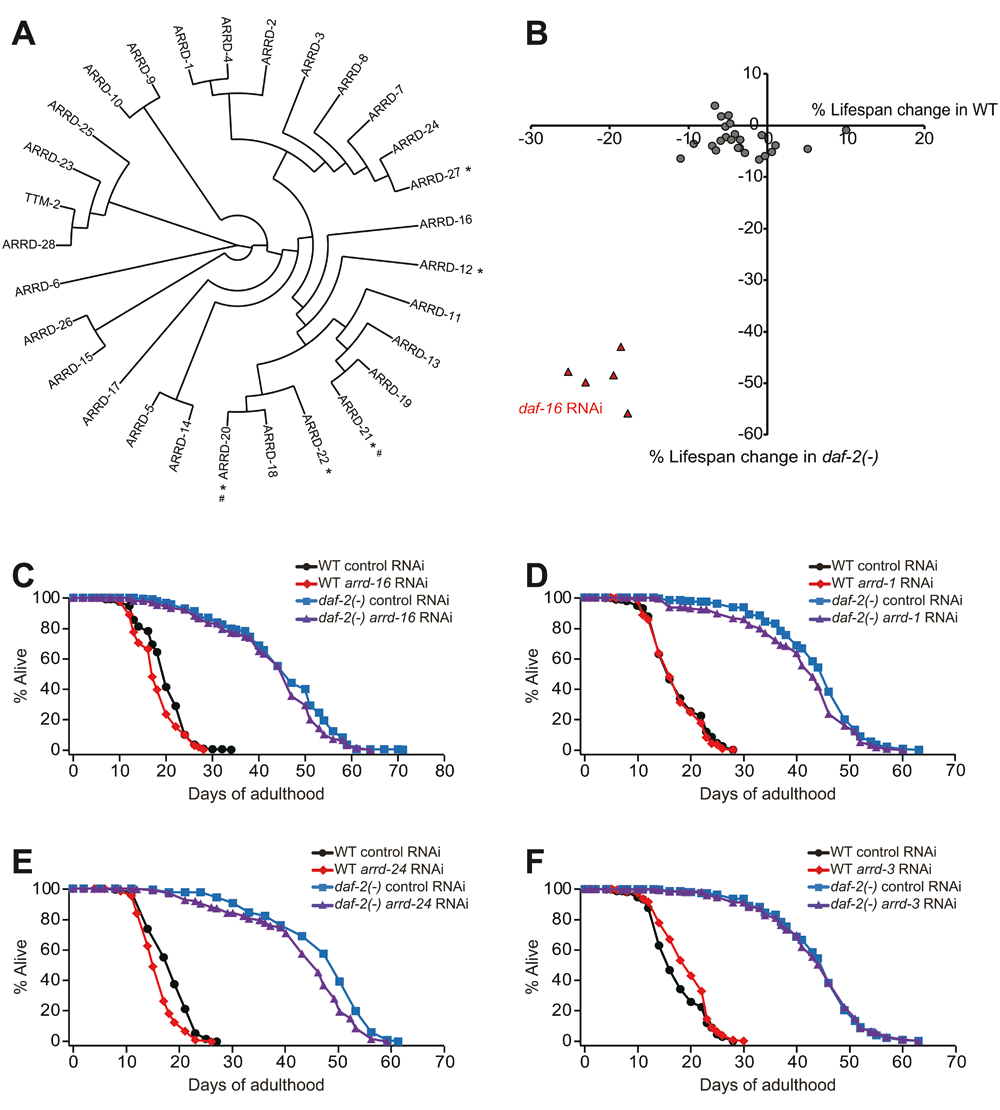
Comments on this article Comments (0)