Keywords
Anastasis, Cell Death, Cell Survival, Cell Suicide, Gene Expression, Recovery, Repair, Reversal of Apoptosis
Anastasis, Cell Death, Cell Survival, Cell Suicide, Gene Expression, Recovery, Repair, Reversal of Apoptosis
Apoptosis (Greek for “falling to death”) was generally assumed to be an irreversible cell suicide process because it involves rapid and massive cell destruction1–7. During apoptosis, intrinsic and extrinsic pro-apoptotic signals can converge at mitochondria, leading to mitochondrial outer membrane permeabilization (MOMP), which releases cell execution factors, such as cytochrome c to trigger activation of apoptotic proteases including caspase-3 and -78,9, small mitochondria-derived activator of caspases (Smac)/direct IAP binding protein with low pI (DIABLO) to eliminate inhibitor of apoptosis protein (IAP) inhibition of caspase activation10,11, and apoptosis-inducing factor (AIF) and endonuclease G to destroy DNA12–15. Activated caspases commit cells to destruction by cleaving hundreds of functional and structural cellular substrates2,16. Crosstalk between signalling pathways amplify the caspase cascade to mediate cell demolition via nucleases (DNA fragmentation factor [DFF]/caspase-activated DNase [CAD]) to further destroy the genome17–19, and alter lipid modifying enzymes to cause membrane blebbing and apoptotic body formation20,21. Therefore, cell death is considered to occur after caspase activation within a few minutes22,23.
However, we and other groups have demonstrated reversal of early stage apoptosis, such as externalization of phosphatidylserine (PS) in cultured primary cells and cancer cell lines24–27. We have further demonstrated that dying cells can reverse apoptosis even after reaching the generally assumed “point of no return”, such as MOMP-mediated cytochrome c release, caspase activation, DNA damage, nuclear fragmentation, and apoptotic body formation26–28. Our observation of apoptosis reversal at late stages is further supported by an independent study, which shows recovery of cells after MOMP29. To detect reversal of apoptosis in live animals, we have further developed a new in vivo caspase biosensor, designated “CaspaseTracker”30, and successfully identified and tracked somatic, germ and stem cells to survive transiently-induced cell death, and potentially during normal development and homeostasis in Drosophila melanogaster after caspase activation30,31, the hallmark of apoptosis2,32. We refer to this recovery phenomenon as “anastasis”27, which means “rising to life” in Greek, for the reversal of apoptosis. Anastasis appears to be an intrinsic cell survival phenomenon, as removal of cell death stimuli is sufficient to allow dying cells to recover26–28,30.
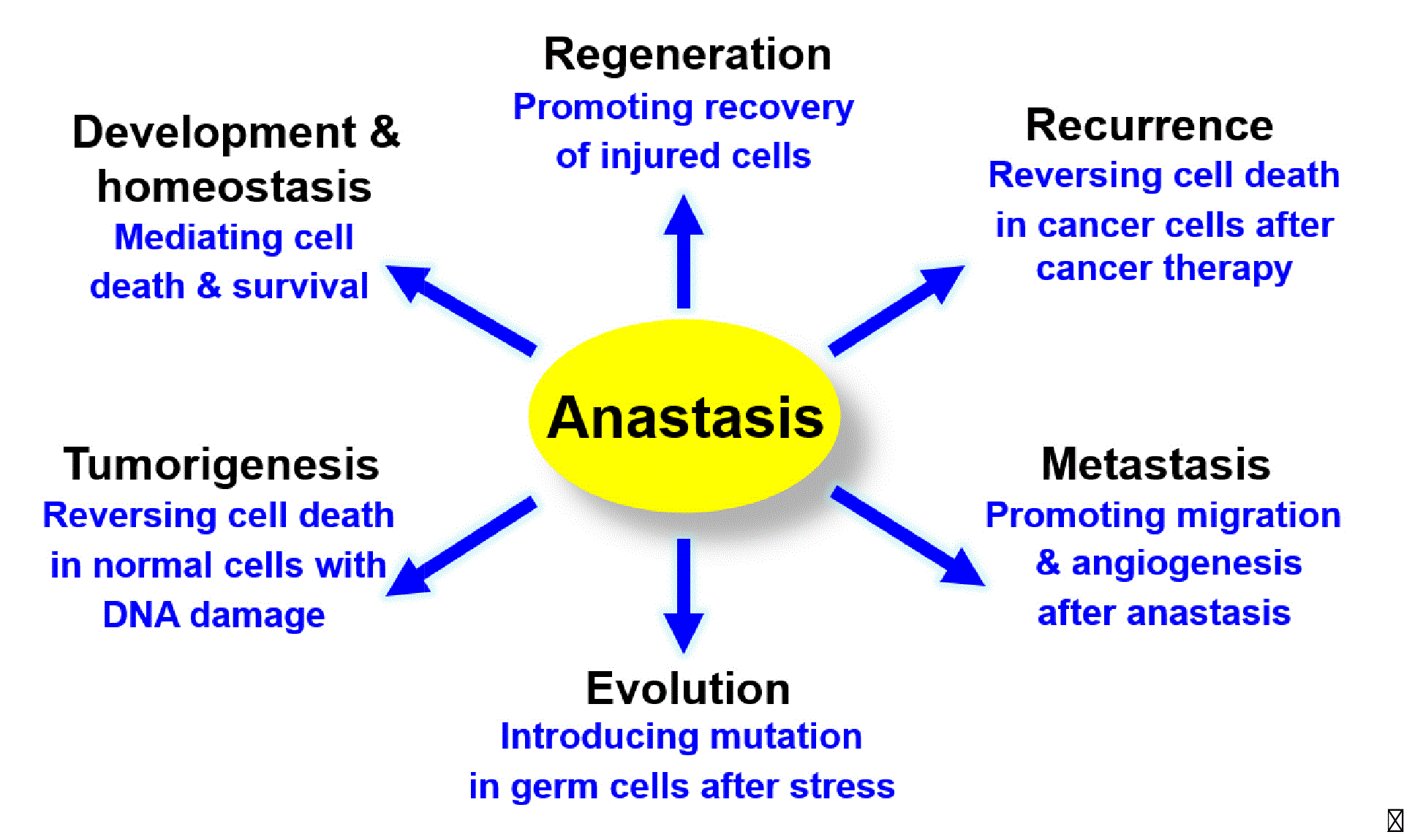
The physiological, pathological and therapeutic importance of anastasis is not yet known. We proposed that anastasis could be an unexpected tactic that cancer cells use to escape cancer therapy26–28. Many tumours undergo dramatic initial responses to cell death-inducing radiation or chemotherapy33–36; however, these cells relapse, and metastasis often occurs in most types of cancer33–35. Therefore, the ability of cells to recover from transient induction of cell death may allow tumour cells to escape treatment, and survive and proliferate, resulting in relapse26–28. Furthermore, cells may acquire new oncogenic mutations and transformation phenotypes during anastasis27,28, such as DNA damage caused by apoptotic nucleases. Therefore, anastasis could be one mechanism underlying the observation that repeated tissue injury increases the risk of cancer in a variety of tissues37, such as liver damage due to alcoholism38, chronic thermal injury in the oesophagus induced by the consumption of very hot beverages39–41, evolution of drug resistance in recurrent cancers26–28, and development of a second cancer during subsequent therapy42–45. Anastasis can also occur in primary cardiac cells and neuronal cell lines27,28, and potentially in cardiomyocytes in vivo following transient ischemia46. These findings suggest anastasis as an unexpected cellular protective mechanism. Therefore, uncovering the mechanisms of anastasis may provide new insights into the regulation of cell death and survival, and harnessing this mechanism via suppression or promotion of anastasis would aid treatment of intractable diseases including cancer, heart failure and neurodegeneration.
Our previous study demonstrated reversibility of ethanol-induced apoptosis at late stages in mouse primary liver cells, and revealed that new transcription is important to reverse apoptosis27,28. During recovery, there was up-regulation of genes involved in pro-survival pathways and DNA damage responses during anastasis (Bag3, Mcl1, Dnajb1, Dnajb9, Hsp90aa1, Hspa1b, and Hspb1, Mdm2)27. Interestingly, inhibiting some of those genes by corresponding specific chemical inhibitors significantly suppresses anastasis27. However, the molecular mechanism of anastasis remains to be elucidated. To study the cellular processes of anastasis, we performed time-course RNA microarray analysis to determine the gene expression profiles of cultured mouse primary liver cells undergoing anastasis following exposure to ethanol, and identified unique gene expression patterns during reversal of apoptosis. Here, we present our time-course microarray data, which reveals the molecular signature of anastasis.
Mouse primary liver cells were isolated from BALB/c mice using collagenase B and cultured as described27,47. The cells were treated with 4.5% ethanol in DMEM/F-12 (DMEM:nutrient mixture F-12) supplemented with 10% fetal bovine serum, 100 U/ml penicillin, and 100 μg/ml streptomycin (Life Technologies, Carlsbad, CA, USA) at 37°C under an atmosphere of 5% CO 2/95% airfor 5 hours (R0), and then washed and further incubated in fresh culture medium for 3 hours (R3), 6 hours (R6), 24 hours (R24), and 48 hours (R48). Three biological replicates were performed at each time point. The untreated cells served as control (Ctrl). Total RNA in the corresponding cell conditions was harvested using TRIzol Reagent, and RNA was purified using the RNeasy Mini Kit (Qiagen, Cologne, Germany). Reverse transcription was performed using SABiosciences C-03 RT2 First Strand Kit to construct cDNA (SABiosciences-Qiagen, Frederick, MD, USA). The cDNA samples were analysed using the Illumina MouseWG-6 v2.0 Expression BeadChip (Illumina, San Diego, CA, USA).
The Partek Genomics Suite 6.6 (Partek, St. Louis, MO, USA) was used for principal component analysis48. Spotfire DecisionSite 9.1.2 (TIBCO, Palo Alto, CA, USA) platform was used to evaluate the fold change of gene expression levels between time points when compared with a common starting point49. Signal values were converted into log2 space and quality control tests were performed to ensure data integrity by comparing the signals of the three biological replicates at each time point. The fold change was based on averaged values of the three replicates at each time point; two-sample Student's t-test was used to determine statistical significance as p-values of less than 0.05, using Partek Genomics Suite v6.5 (Partek Inc., St. Louis, MO, USA).
For the time-course gene expression analysis using Spotfire, all time points were compared with time point Ctrl, which represents untreated cells. Spotfire was used to show the genes that displayed specific changes in gene expression after removal of apoptotic inducers for 3 hours and 6 hours, as well as the genes that were up-regulated from apoptosis (R0) to 6 hours (R6) after removal of the inducer. Genes with specific and significant change (Log2 > 1 or <-1) in expression at the corresponding timepoint are highlighted. Interaction network analysis of the up-regulated genes during anastasis was performed using GeneMANIA database (http://genemania.org/)50,51.
We have demonstrated that mouse primary liver cells can reverse the apoptotic process at the execution stage, despite experiencing important checkpoints commonly believed to be the “point of no return”, including caspase-3 activation, DNA damage, and cell shrinage27,28. To pursue the mechanisms of anastasis, we performed time-course high-throughput microarray to evaluate gene expression profiles during reversal of ethanol-induced apoptosis in mouse primary liver cells. RNA samples were collected from the untreated primary liver cells (Ctrl), the cells treated with 4.5% ethanol for 5 hours when cells exhibited hallmarks of apoptosis (R0), and the treated cells that were then washed and cultured in fresh medium for 3 (R3), 6 (R6), 24 (R24) and 48 (R48) hours. Apoptosis was confirmed previously in the ethanol-treated cells (R0), which displaced hallmarks of apoptosis, including plasma membrane blebbing, cell shrinkage, cleavage of caspase-3 and its substrates, such as PARP and ICAP (Figure 1A and B, images reprinted with permission27). The features of apoptosis vanished after removal of the cell death inducer (R24), indicating recovery of the cells (Figure 1A and B). Three biological replicates were performed at each time point. Principal component analysis indicated that all three biological replicates of each time point exhibited a very high correlation, as indicated by clustering, for the dataset of all 18 samples (Figure 2A; see Data availability52). Unsupervised hierarchical clustering confirms the similarity between all the replicates at each time point (Figure 2B; see Data availability52; Supplementary Figure 1).
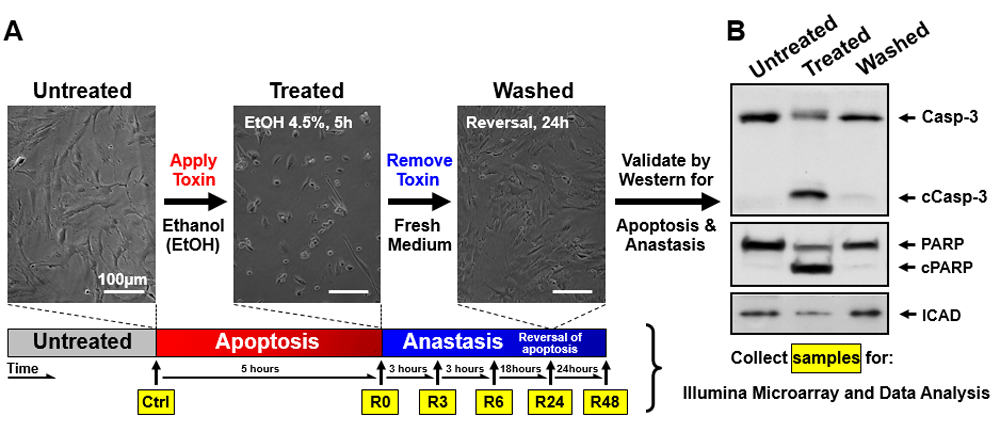
Mouse primary liver cells were treated with 4.5% ethanol for 5 hours (R0) and then washed and cultured in fresh medium for 3 (R3), 6 (R6), 24 (R24), and 48 (R48) hours. The untreated cells served as control (Ctrl). (A) Light microscopy and (B) western blot analysis validated apoptosis to occur at R0, and anastasis at R24. Cells were collected at the indicated timepoints of (A) for RNA extraction. Gene expression profiling was performed by microarray, and analysed by Spotfire. The images from Figure 1A and B are adopted from the Mol Biol Cell 23, 2240–52 (2012)27. Reprinted with permission.
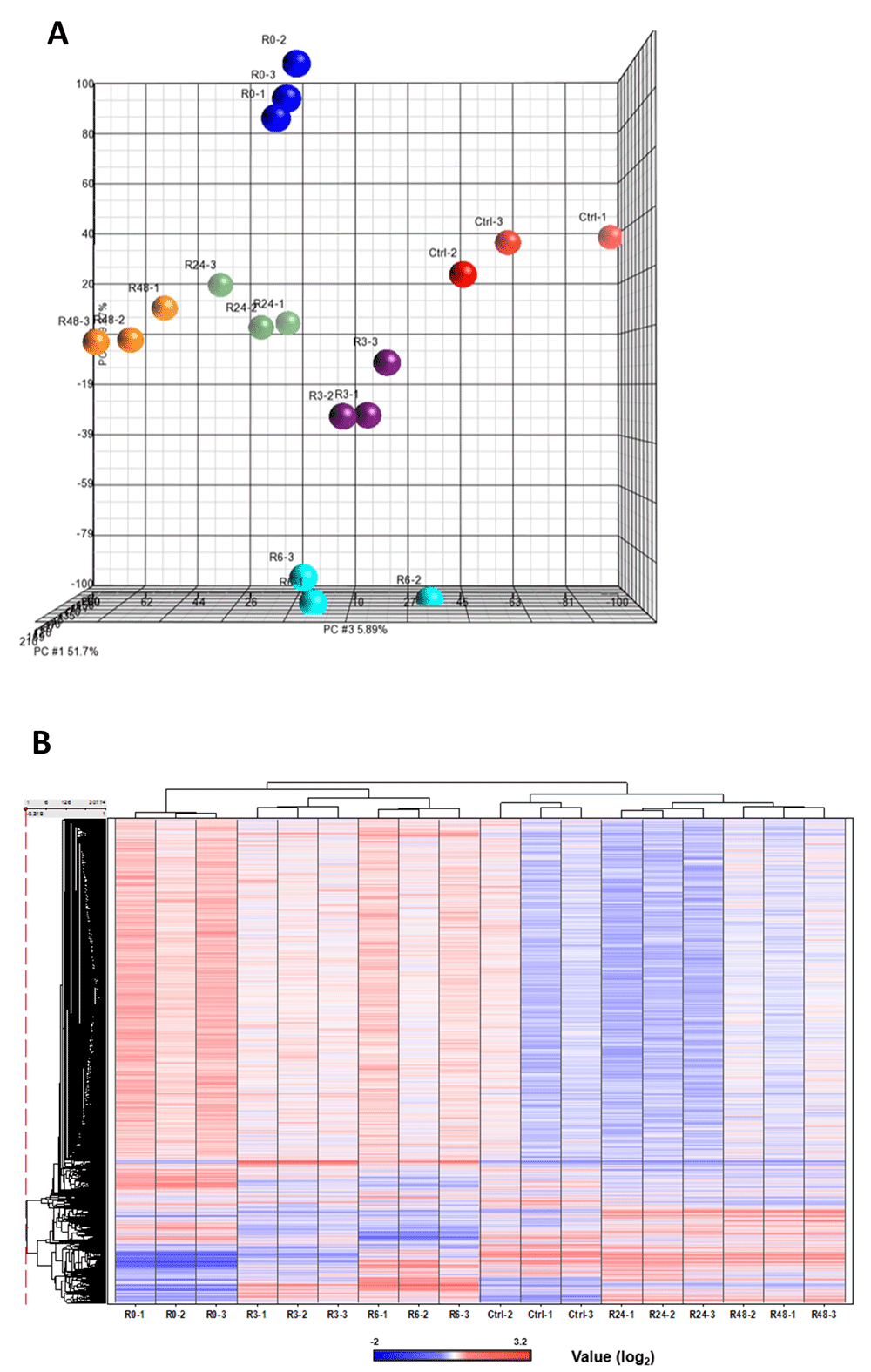
The three biological replicate samples of microarray data were shown to cluster together by using (A) principal component analysis (PCA) and (B) unsupervised hierarchical clustering of the RNA microarray data of eighteen samples.
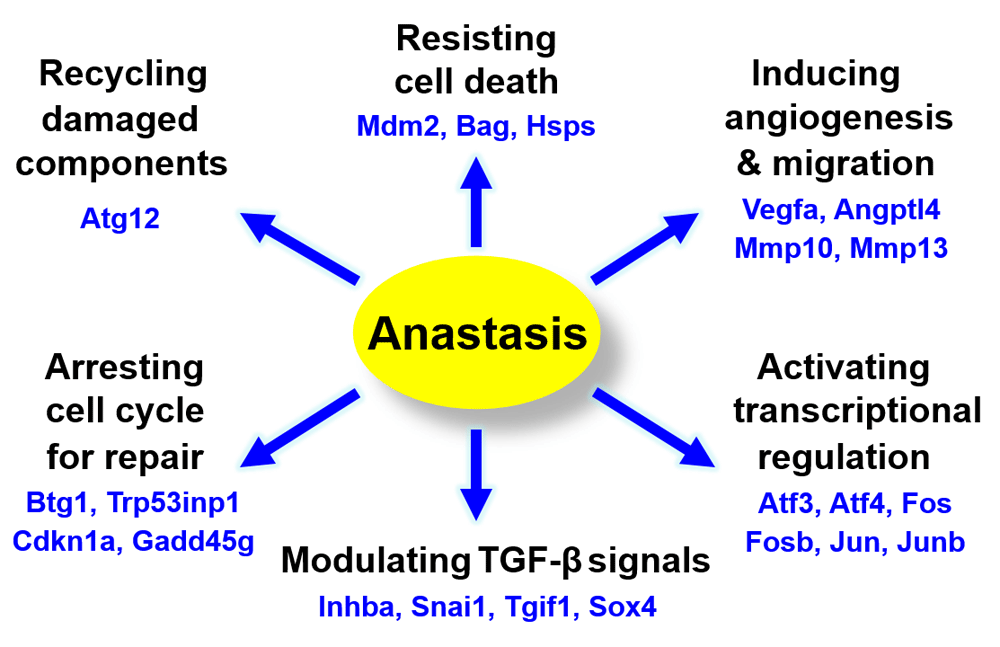
Genes that display significant changes in expression during anastasis at the earliest time point of 3 hours, following the removal of the apoptotic inducer, may represent critical first responders of anastasis (Figure 3A, Table 1), including transcription factors of the activator protein-1 (AP-1) family (Atf3, Fos, Fosb, Jun, Junb), transforming growth factor‑β (TGF-β) signal pathway and its related regulators (Inhba, Snai1, Tgif1, Sox4, Klf4, Klf6, Klf9), pro-survival Bcl-2 family member (Bag3), inhibitor of p53 (Mdm2), anti-proliferation (Btg1), DNA damage (Ddit3, Ddit4) and stress-inducible (Dnajb1, Dnajb9, Herpud1, Hspb1, Hspa1b) responses. Starting at 6 hours of anastasis, other groups of gene pathways displace the peak of transcription, such as cell cycle arrest (Cdkn1a, Trp53inp1), autophagy (Atg12, Vps37b), and cell migration (Mmp10 and Mmp13) (Figure 3B, Table 2 and Table 3). Expression of potent angiogenic factors, such as Vegfa and Angptl4, peaks at 3 and 6 hours of anastasis, respectively. Changes in expression of most of these genes peak at the 3–6-hour time points after removal of the apoptotic stimulus and then return to baseline (Figure 3A and B; Supplementary Figure 1). Interestingly, certain genes involve in splicing of pre-mRNA (Rnu6), and growth arrest and DNA repair (Gadd45g) stay up-regulated during both apoptosis and anastasis (Figure 3C; Supplementary Figure 1).
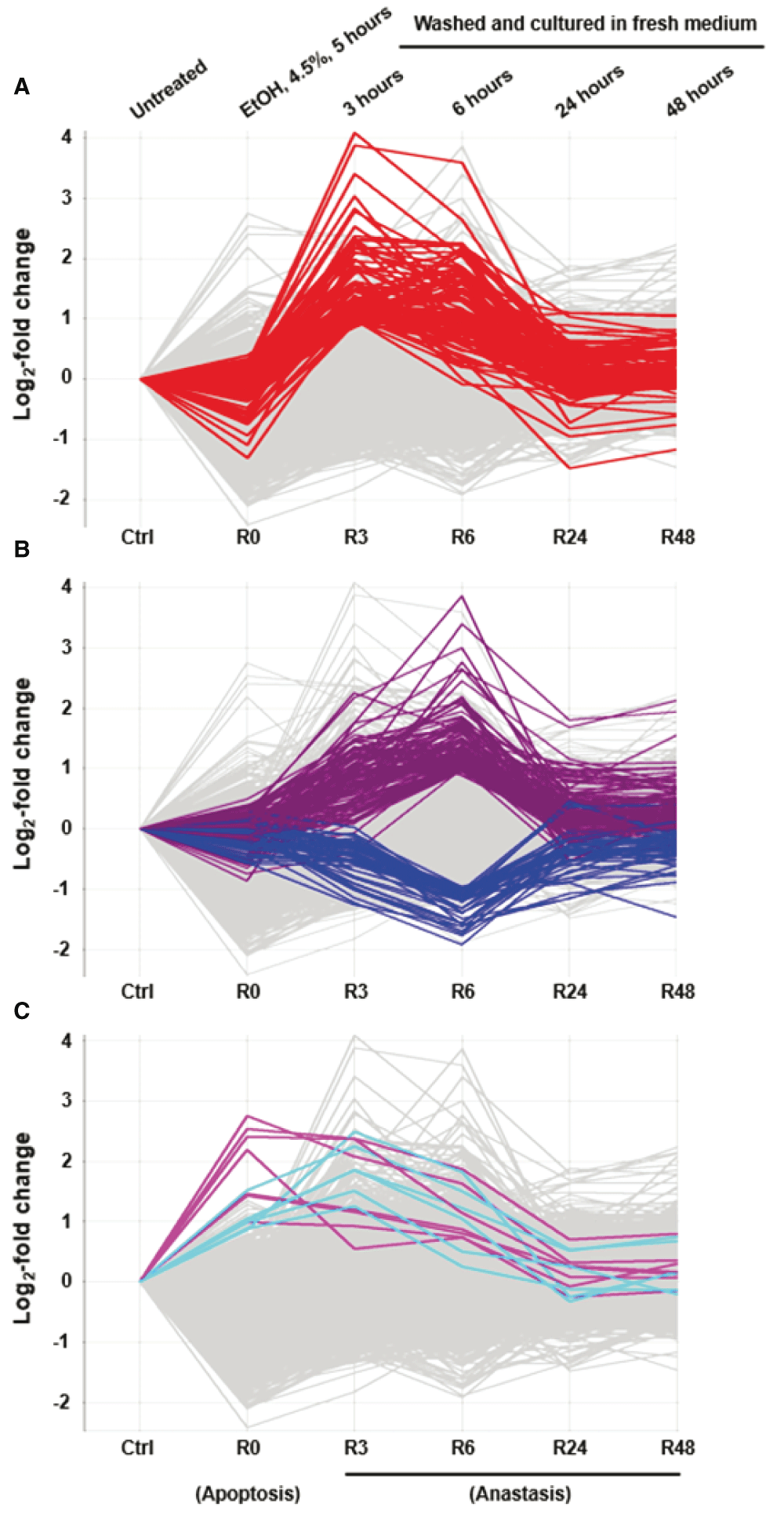
Log2 fold change of gene expression comparison between untreated cells (Ctrl), ethanol-induced apoptotic cells (R0), and induced cells that were then washed and further cultured in fresh medium for 3 (R3), 6 (R6), 24 (R24), and 48 (R48) hours. Genes that displaced specific (A) up-regulation at R3, (B) up- or down-regulation, and (C) up-regulation during R0 to R6 absolute log2 fold change >1 are highlighted. The log2 signal values from three biological replicates were averaged (geometric mean) for each time point.
The change in transcriptional profiles during anastasis provides us mechanistic insights into how dying cells could reverse apoptosis (Figure 4). In early anastasis (R3), our microarray data reveals that the regulators of the TGF-β signalling pathway, which control various fundamental cellular process, including proliferation, cell survival, apoptosis and transformation53–55, are upregulated. The activation of the TGF-β pathway is further supported by the upregulation of AP-1 (Jun-Fos) during early anastasis. The up-regulation of the TGF-β pathway also promote the expression of murine double minute 2 (Mdm2)56,57, an inhibitor of p53 that is also up-regulated during early anastasis27. As p53 plays a critical role in regulating apoptosis and DNA repair58,59, the expression of Mdm2 could not only promote cell survival by inhibiting p53-mediated cell death, but also cause mutations as we have observed in the cells after anastasis27. Expression of Mdm2 can also activate XIAP60, which inhibits caspases 3, 7 and 961–66, and therefore, could promote anastasis by suppressing the caspase-mediated cell destruction process. Up-regulation of anti-apoptotic BCL2 protein (Bag3) and heat shock proteins (Hsps) during anastasis can also neutralize pro-apoptotic proteins to promote cell recovery67–69. Notably, Bbc3 is a pro-apoptotic BH3-only gene to encode PUMA (p53 upregulated modulator of apoptosis)70,71. Its expression peaks at anastasis (R3-R6), suggesting the sign of anastasis vs apoptosis in the recovering cells during the early stage of the cell recovery process.
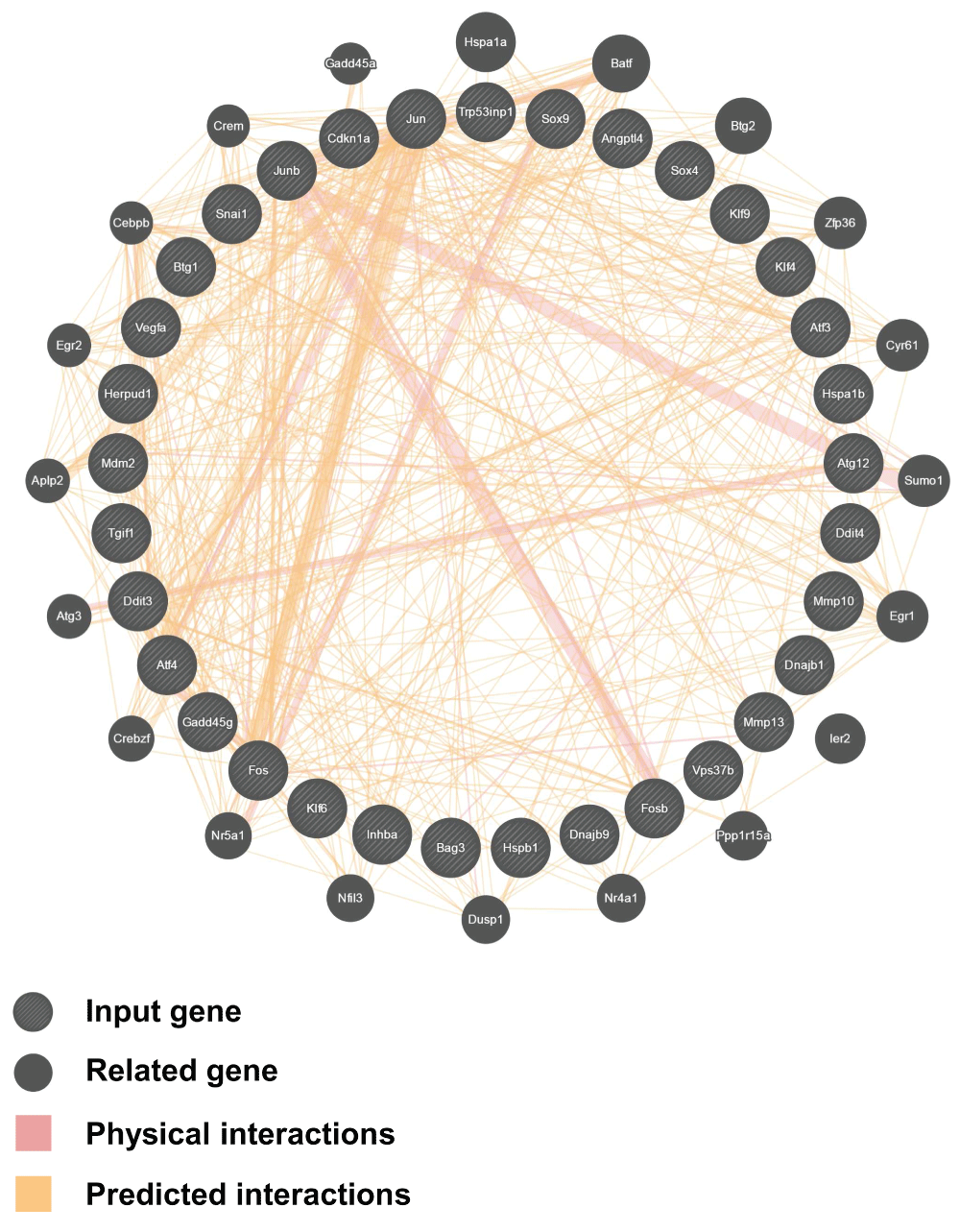
The 33 up-regulated genes during anastasis were selected for analysis using GeneMANIA.
To reverse apoptosis, the recovering cells need to remove or recycle the destroyed cellular components, such as the toxic or damaged proteins that are cleaved by caspases, and dysfunctional organelles like the permeabilized mitochondria. Autophagy could contribute to anastasis, as the recovering cells display up-regulation of Atg12 (Figure 3B, Table 2), which is important to the formation of autophagosome to engulf the materials that are then transported to lysosomes or vacuoles for degradation72–75. Recently studies reveal that autophagy can be activated by the DNA damage response, and play a role in maintaining the nuclear and mitochondrial genomic integrity through DNA repair and removal of micronuclei and damaged nuclear parts76,77. This could suppress mutagenesis and oncogenic transformation to occur in the cells that reverse apoptosis as observed after DNA damage27,28. Autophagy is also implicated in the exosome secretory pathway78–80, which could allow rapid clearance of damaged or toxic materials during anastasis through exosomes. Interestingly, our microarray data shows that the recovering cells display up-regulation of potent angiogenic factors such as Vegfa and Angptl4 (Figure 3A and B, Table 1 and Table 2), which promote vascular permeability and angiogenesis81–84. This could facilitate anastasis by supplying nutrient and clearing waste products. However, this could also enhance tumour progression and metastasis when anastasis occurs cancer cells. In fact, our data also reveals the up-regulation of genes involved in cell migration during anastasis27, such as Mmp 10 and 13 that encode matrix metalloproteinases85–88. This could be a stress-inducible response that promotes cell migration, like what was observed in HeLa cells after anastasis28, which might contribute to wound healing, or metastasis during cancer recurrence89,90.
Arresting cell cycle during anastasis is important as it can allow damaged cells to be repaired before they restore proliferation. This hypothesis is supported by the microarray data that reveals up-regulation of genes that suppress cell cycle (Figure 3A–C). For example, B-cell translocation gene 1 (Btg1) is an anti-proliferative gene91,92, which is up-regulated during the early anastasis (R3). At later stage of anastasis (R6), other cell cycle inhibitors express, including Cdkn1a which encodes p21 that induces cell cycle arrest and senescence93–95, and also Trp53inp1 which encodes tumor protein p53-inducible nuclear protein 1 that can arrest cycle independent to p53 expression96. These suggest that cell cycle is suppressed by multiple pathways during anastasis.
We also identified genes that are up-regulated both during apoptosis and anastasis, such as Gadd45g, and Rnu6 (Figure 3C, Table 4). Gadd45g functions in growth arrest and DNA repair97,98, and therefore, could be the cytoprotective mechanism that preserves the dying cells during cell death induction (R0), and promotes the injured cells to repair when environment is improved (R3 and R6). Rnu6 encodes U6 small nuclear RNA, which is important for splicing of a mammalian pre-mRNA99–102. Upregulation of Rnu6 from R0 to R6 suggests that post-transcription regulation could involve during apoptosis and anastasis. In fact, translational regulation also contributes to anastasis. For example, caspase-3, PARP and ICAP are cleaved in dying cells during apoptosis, and the non-cleaved form of corresponding proteins restores after anastasis (Figure 1B). Interestingly, the mRNA level of caspase-3 and PARP did not show significant increase (see Data availability52). This suggests translational regulation to occur during anastasis.
Our study provides new insights into the mechanisms and consequences of anastasis (Figure 5) Researchers can analyse our microarray data to further identify the hallmarks of anastasis, understand its role, elucidate molecular mechanisms that reverse apoptosis, and develop therapeutic strategies to control anastasis. To identify the genes that displace specific change on a transcriptional level, software such as Spotfire can be used to view the gene expression pattern at different time points during the reversal of apoptosis49. To study the molecular mechanism of anastasis, Ingenuity Pathway Analysis can be used to create mechanistic hypotheses according to the transcriptional profile103. To identify drugs that modulate anastasis, Connectivity Map can be used to identify small molecules that promote or suppress anastasis based on its gene expression signature104,105. Anastasis could be a cell survival phenomenon mediated by multiple pathways26–28,30, so by comparing the gene expression profiles, researchers can study its potential connection to other cellular processes, such as anti-apoptotic pathways, autophagy, and stress-inducible responses75,106–110. By searching the molecular signature of anastasis, researchers can study the potential contribution to physiopathological conditions, such as metastasis during cancer recurrence, recovery from heart failure and wound healing89,90,111. Further data analysis will stimulate generation of hypotheses for future studies involving anastasis. As our understanding of anastasis mechanism expands, it will uncover its potential impacts on physiology and pathology, and offer exciting new therapeutic opportunities to intractable diseases by mediating cell death and survival (Figure 6).
Raw data for Tang et al., 2016 “Molecular signature of anastasis for reversal of apoptosis” available at: doi, 10.6084/m9.figshare.450273252
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)