Keywords
lifespan, human lifespan, contradictory findings, ageing, life history, refutation
This article is included in the Preclinical Reproducibility and Robustness gateway.
lifespan, human lifespan, contradictory findings, ageing, life history, refutation
We have conducted minor changes to the phrasing of the manuscript regarding projections of MSA and the expected increase in MRAD, and corrected the spelling of "Calment". We have also added Supplementary Figure S1 and Supplementary Figure S2 with reference in the text, initially embedded in the supplementary material code, to address the concerns and comments of reviewers.
See the authors' detailed response to the review by Jean-Michel Gaillard
See the authors' detailed response to the review by Michael R. Rose and Laurence D Mueller
Recent findings by Dong et al.1 suggested fixed upper limits to the human life span. Using the same data, we replicated their analysis to obtain an entirely different result: the upper limit of human life is rapidly increasing.
Dong et al. conclude that the maximum reported age at death (MRAD) is limited to 125 years in humans1 and that lifespan increases above age 110 are highly unlikely, due to the reduced rate of increase in life expectancy at advanced ages.
We repeated Dong et al.’s1 analysis using identical data (SI). Replicating these findings requires the inclusion of rounding errors, treating zero-rounded values as log(1) and the incorrect pooling of populations.
The Human Mortality Database (HMD) data provide both the age-specific probability of survival (qx) and the survival rates of a hypothetical cohort of 100,000 individuals (lx). However, lx survival rates are rounded off to the nearest integer value.
The magnitude and frequency of lx rounding errors increases as the probability of survival approaches 1 in 100,000. These rounding errors mask variation in survival rates at advanced ages: over half of lx survival data are rounded to zero above age 90 (Figure 1b).
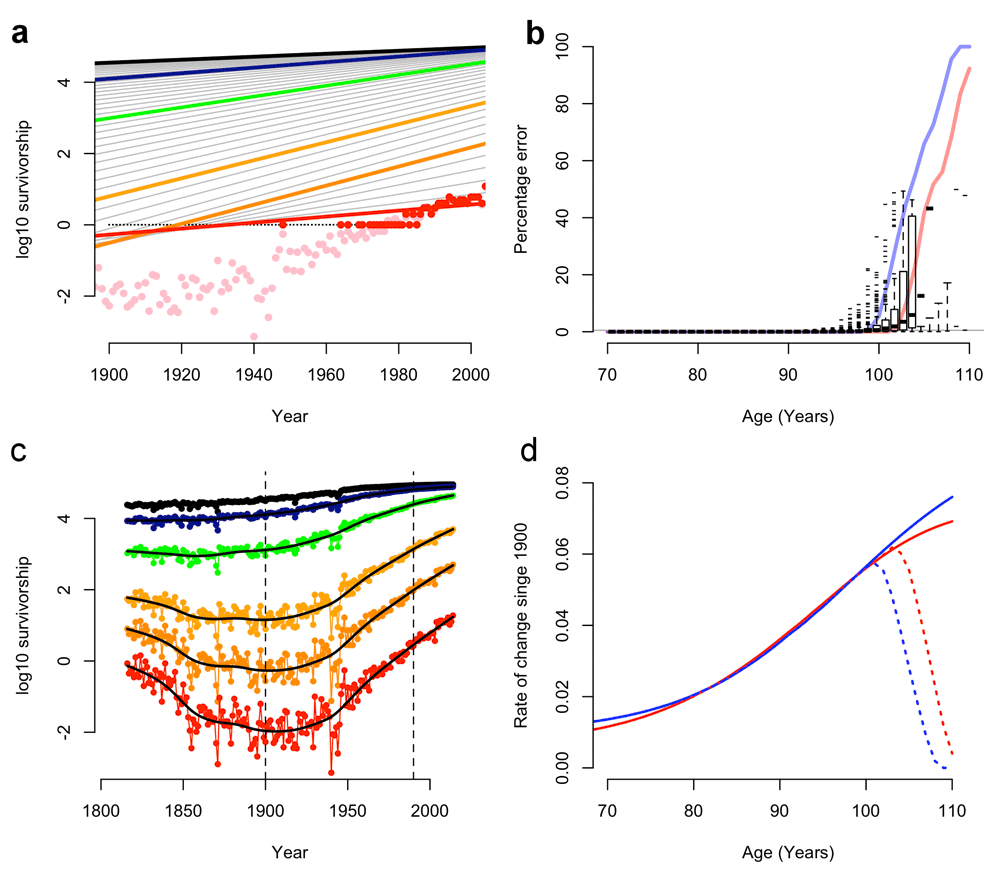
(a) Figure modified after Dong et al. Figure 1b, showing rounded survival data (red points), rounded survival data with log(0)=log(1) (black points), the resulting linear regression in Dong et al. (solid red line) and observed survival data (pink points). (b) Rounding errors in survival data (box-whisker plots; 95% CI) and the proportion of survival data rounded to zero in males (blue line) and females (red line). (c) Survival data from (a) with rounding errors removed, showing variation outside the 1900-1990 period (vertical dotted lines). (d) The rate of change in late-life mortality since 1900 with (dotted lines) and without (solid lines) rounding errors (after Dong et al. Figure 1c).
Dong et al. appear to have used these rounded-off survival data in their models1 and incorrectly treated log(0) values as log(1) in log-linear regressions (Figure 1a–d; SI).
These errors have considerable impact. Re-calculating cohort survival from raw data or excluding zero-rounded figures eliminates the proposed decline in old-age survival gains (Figure 1d; SI).
Likewise, recalculating these data removed their proposed limits to the age of greatest survival gain (SI), which in 15% of cases were the result of the artificial 110-year age limit placed on HMD data2.
We also found that variation in the probability of death was masked by date censoring1. Major non-linear shifts in old-age survival occur outside the 1900–1990 period used by Dong et al. (Figure 1c). Why these data were excluded from this regression, but included elsewhere, is unclear.
Evidence based on observed survival above age 110 appears to support a late-life deceleration in survival gains1. For the period 1960–2005 Dong et al. present data1 from 4 of the 15 countries in the International Database on Longevity3 (IDL). In their pooled sample of these countries, there is a non-significant (p=0.3) reduction in MRAD between 1995 and 2006 (Figure 2a).
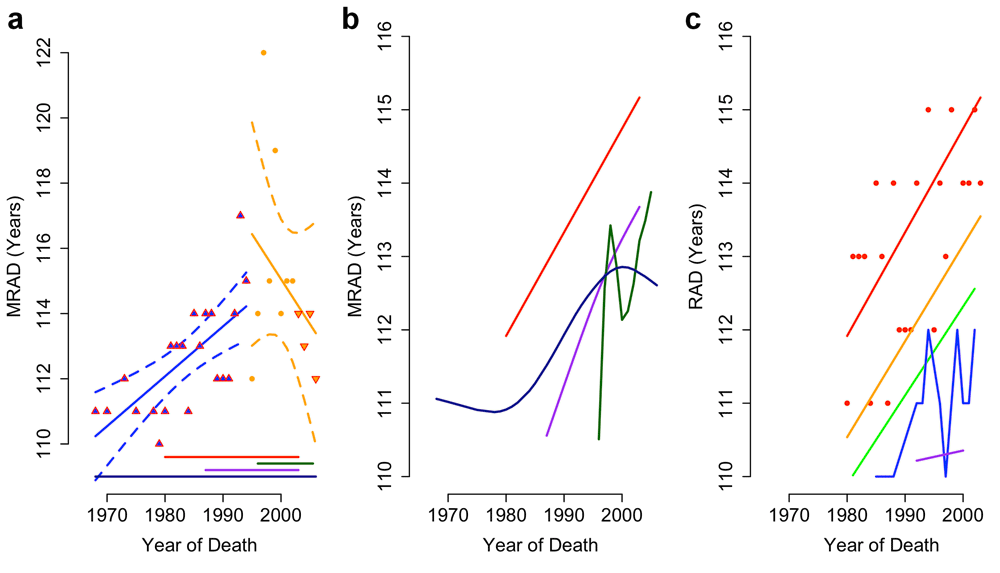
(a) Reproduction of Dong et al. Figure 2a, including 95% CI for increasing (p<0.0001) and falling (p=0.3) maximum recorded age at death (MRAD), showing data biased by the addition and removal (up and down arrows) of populations. (b) Locally weighted smoothed splines of MRAD in Japan (green), the USA (red), the UK (dark blue) and France (purple). (c) Locally weighted trends of MRAD in the USA across the oldest 5 reported ages at death (red, orange, green, blue and purple lines show rank 1–5 respectively).
The declining MRAD reported by Dong et al.1 arises from the use of falling sample sizes. According to the Gerontology Research Group (GRG), 62% of validated supercentenarians alive in 2007 resided in France and the USA. However, these countries are not surveyed3 by the IDL after 2003 (Figure 2a). The proposed post-1995 decline in MRAD results from this dramatic fall in sample size.
Viewed individually, all four countries have an upward trend in the mean reported age at death (RAD; Figure 2b) of supercentenarians (SI) and the top 5 ranked RADs (Figure 2c). All four countries achieved record lifespans since 1995, as did 80% of the countries in the IDL. Without the pooling of IDL data used by Dong et al. there is no evidence for a plateau in late-life survival gains.
We attempted to reproduce Dong et al.’s supporting analysis of GRG records. The text and Extended Data Figure 6 of Dong et al. do not match annual MRAD records from 1972 as stated1. However, they do match deaths of the world’s oldest person titleholders from 1955 (GRG Table C, Revision 9) with all deaths in May and June removed (SI).
Actual MRAD data from the GRG support a significant decline in the top-ranked age at death since 1995 (r = -0.47; p = 0.03, MSE = 3.2). However, this trend is not significant if only Jeanne Calment is removed (p = 0.9). Linear models fit to lower-ranked RADs have an order of magnitude better fit, and all indicate an increase in maximum lifespan since 1995 (Supplementary Figure S1; N= 64; SI).
Collectively, these data indicate an ongoing rebound of upper lifespan limits since 1950, with a progressive increase in observed upper limit of human life. To estimate theoretical limits, we developed a simple approximation of the upper limit of human life.
Mortality rates double with age in human populations (Figure 3a and b). Log-linear models fit to this rate-increase closely approximate the observed age-specific probability of death4. These models also provide a simple method of predicting upper limits to human life span that is independent of population size.
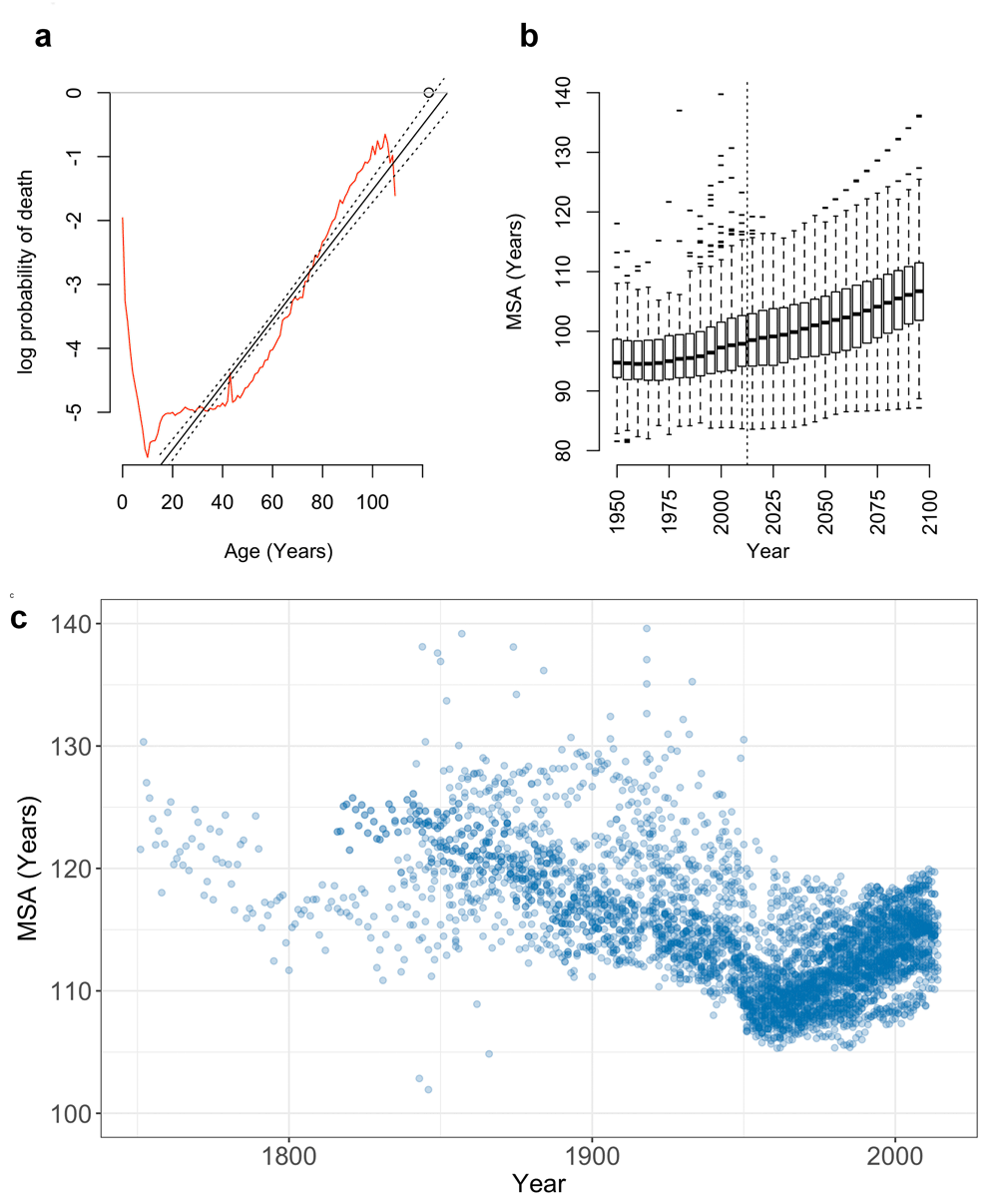
(a) In humans, the probability of death q at age x (qx; red line) increases at an approximately log-linear rate with age (black lines; 95% CI), shown here for the birth cohort of Jeanne Calment (d.122.5 years; circle). Projection of this log-linear increase to log(q) = 0 provides the MSA, the upper limit of human survival, shown here for (b) observed and projected global populations5 and (c) 40 historic HMD populations 1751–2014.
We fit log-linear models to age-specific mortality rates from the HMD data used by Dong et al.1, and used these models to predict the age at which the probability of death intercepts one. This maximum survivable age (MSA) provides a simple, conservative estimate of the upper limit of human life (Figure 3c).
Log-linear models closely approximate the observed probability of death in HMD populations for both period and cohort life tables (median R2 = 0.99; 4501 population-years). The MSA limit is compatible with observed ages at death in the GRG database with 330 out of 331 supercentenarians approaching, but not exceeding, their predicted lifespan limit (Supplementary Figure S2; SI). These models predict an MSA exceeding 125 years within observed historic periods (Figure 3b and c; SI).
Furthermore, period HMD data indicate that MSA is steadily increasing from a historic low c.1956 (Figure 3b and c), tracking the increasing MRAD during the same period. The United Nations5 global mortality data from 194 nations support this trend, with projected population data from the UN predicting a gradual rise in MRAD and MSA through the year 2100 (Figure 3b).
This analysis provides an estimate of human lifespan limits that is conservatively low. Log-linear mortality models assume no late-life deceleration in mortality rates6, which, if present, would increase the upper limits of human lifespan7. In addition, these models are fit to population rates and cannot provide an estimate of individual variation in the rate of mortality acceleration.
Given historical flexibility in lifespan limits and the possibility of late-life mortality deceleration in humans8, these models should, however, be treated with caution.
A claim might be made for a general, higher 130-year bound to the human lifespan. However, an even higher limit is possible and should not be ruled out simply because it exceeds observed historical limits.
Life table data were downloaded from the United Nations5 (UN) and the Human Mortality Database (HMD) and lifespan records from the International Database on Longevity (IDL) and the Gerontology Research Group (GRG).
Least squared linear models were fit to life table data on the log-transformed age-specific probability of death (qx), and projected to qx=1 to predict the maximum survivable age in each population (Figure 1b and c; SI). Maximum lifespan within GRG and IDL data was annually aggregated and fit by locally weighted smoothed splines9 (Figure 3b and c).
We reproduced the analysis of Dong et al. in R version10 3.2.1 (SI), using the code in Supplementary File 1.
An earlier version of this article can be found on bioRxiv (doi: 10.1101/124800).
The authors declare that all data are available within the paper and its supplementary material.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
{kind=link}
Your assertion that our use of the 1900-1990 census period for figure 1c invalidates our findings is false. We have already calculated your regressions using data from 1900-present. We ... Continue reading Dear Sirs,
Your assertion that our use of the 1900-1990 census period for figure 1c invalidates our findings is false. We have already calculated your regressions using data from 1900-present. We have posted the near-identical and highly error prone figure that results from using these dates as a supplementary figure.
The statement that your errors are “greatly overstated by the omission of all data since 1990” is demonstrably false. The error rate of zero-rounded data from 1900-1990 is lower than the error rate of the 1900-2014 data. The magnitude of your errors are not “greatly overstated” in our analysis, they are understated by 16%.
The statement that your data were truncated at 1990 was based on our attempts to reproduce your figures in the absence of any code. This was only possible for figure 1c if we performed a 1900-1990 censoring. Other analysis did not use this census period, and we did not use this census period throughout our paper.
Furthermore, we have previously used data from 1900-present, without the 1990 censoring, for all analyses. We found no difference in the outcome or the scope of your errors when using this data, except that use of 1900-1990 data reduces the error rate and better reproduces fig. 1c.
Therefore, the 1900-1990 truncation had no material effect on our analysis. All of our data was also calculated from 1900-present, 1925-present, from 1850-present, from 1800-present, from 1950-present, and across many other time periods, in the hope that we may arrive at a trend that supported your original findings.
Not one of these periods illustrated any plateau in MRAD.
There are several other points that you have misconstrued. The first is that the reasoned criticisms of JMG support your findings, despite his statement that “I am quite convinced by all the problems the authors identified in the analysis performed by Dong et al.”. JMG has provided an insightful comment on our methodology, which we believe results from a simple misunderstanding and has no bearing on the errors in your paper.
The "improperly formed" boxplots of error rates reflect the calculated maximum, median, 95th percentile and 75th percentile of error rates all reaching 100%. That is, the frequency of your zero-rounding errors becomes so high, and error rates below 100% are so rare, there is no longer a 'box' forming in these boxplots.
This was the rationale for including the red and blue lines, showing the overall fraction of data rounded to zero.
As for your comments on the IDL and GRG data, we would highlight several problems. First, you cannot ignore your incorrect pooling of the IDL data because of trends in another database.
There is no evidence that the proposed plateau in MRAD occurs in any single one of these populations. The ‘plateau’ trend that appears when you pool these data is, as we have shown in our code, a result of poor assumptions. You cannot pool four populations in this way, when two of them are only present to 2003 and one to 2006. You have done nothing to address these problems, other than repeating your initial mistake.
Your ‘obvious’ plateau in the GRG is, as we have stated before, based on a single data point. Removal of that data point removes your proposed plateau in these data. It also makes a simple increasing linear model, with no post-1995 plateau in MRAD, the best fit for observed data under the R squared metric, the Bayesian information criteria and the Akaike information criteria.
We find it remarkable that a selected-by-eye regression, with a worse fit than a linear model when Jeanne Calment is removed, is a core supporting argument for your paper.
As we highlighted for JMG, before Jeanne Calment the record-holder for lifespan, Mary Beard held this record. Mary Beard’s record was retracted after being considered valid for 24 years, when it was recognized her extreme age was a straightforward clerical error. As you admit, a similar clerical error for Jeanne Calment would undo your proposal for a significant fall in MRAD.
The GRG acknowledge this inherent unreliability, stating quite clearly that their data are not complete surveys of the oldest old. They also state that many supercentenarian records are subsequently found to be false positives, or are impossible to verify. Therefore, the analysis presented in Nature depends on a single data point, drawn from unreliable data, gathered from incomplete surveys with a high false positive rate, conducted in a small number of countries.
I suggest you ask a statistician whether they consider it good practice to extract species-wide trends from such data.
In contrast, we have based our analysis on surveys of global populations, conducted by national statistical bodies. None of our calculated limits depend on GRG or IDL data.
Your comment that the median MSA we observed, of around 100 years, supports your results is unfounded. That MSA reflects diversity across all countries in the UN database. It better reflects the limit of lifespan in Namibia than in France, or in any of the developed countries you measured. Furthermore, this median is increasing linearly. Like almost all global populations, the MSA is increasing, with no sign of a plateau.
We have provided data on the correspondence between our MSA estimates and observed data: of 311 observations, 310 were consistent with limits calculated under MSA (our figure S2).
Your suggestion that these data are implausible is not based on any apparent analysis. You have not provided any indication why these estimates are incorrect, beyond your own opinion. Where is your code?
We would also highlight that there is nothing historically remarkable about the current length of the Jeanne Calment's lifespan record. We have heard the argument that holding an unbroken lifespan record for two decades somehow suggests an MRAD plateau, an argument you repeated in response to Rozing, Kirkwood and Westendorp (Nature 546, E12 (29 June 2017) doi:10.1038/nature22789).
However, the overall lifespan record was previously held for 17 consecutive years (Mary Kelly, 1964-1981). The male lifespan record was held for 68 consecutive years (Gert Adrians-Boomgaard) and then 21.9 years (Mathew Beard 1985-2007) by a margin of 3 years. None of these broken records indicated a plateau in MRAD.
Neither does the lifespan record of Jeanne Calment. We suggest the Jeanne Calment record simply reflects the uncertain ascertainment, and rarity of adequate documentation, required to validate the oldest-old.
Finally, I consider the findings of Dong et al. are invalid as a result of methodological errors, errors you have not corrected or addressed. Pointing to data you feel supports your view, in this paper or others, does not answer any of the problems raised by ourselves or other authors.
Were our results to entirely support your proposed MRAD plateau this would do nothing to excuse or explain your errors in analysis.
I cannot speak for my (currently traveling) co-author, but I find it disturbing that you have failed to acknowledge or correct even the most basic errors in typesetting and analysis, even those that have little material effect on the result (your Extended Data figure 6 still has incorrect axes, is not the data you said it was in the text, and is still missing all the people who died in May and June).
It seems to many observers that you have committed basic errors that you refuse to acknowledge or repair.
Despite our reproduction of your analysis, you have not provided a single piece of code. We have repeatedly requested this code. Should you ever provide it, in accordance with the editorial guidelines of Nature, I will gladly highlight these errors for you.
In the mean time, I suggest that you reflect on the possibility of honest errors driving your results.
Regards,
Saul Newman
https://f1000researchdata.s3.amazonaws.com/linked/166727.Fig_1_HiRes_2014.tiff
Your assertion that our use of the 1900-1990 census period for figure 1c invalidates our findings is false. We have already calculated your regressions using data from 1900-present. We have posted the near-identical and highly error prone figure that results from using these dates as a supplementary figure.
The statement that your errors are “greatly overstated by the omission of all data since 1990” is demonstrably false. The error rate of zero-rounded data from 1900-1990 is lower than the error rate of the 1900-2014 data. The magnitude of your errors are not “greatly overstated” in our analysis, they are understated by 16%.
The statement that your data were truncated at 1990 was based on our attempts to reproduce your figures in the absence of any code. This was only possible for figure 1c if we performed a 1900-1990 censoring. Other analysis did not use this census period, and we did not use this census period throughout our paper.
Furthermore, we have previously used data from 1900-present, without the 1990 censoring, for all analyses. We found no difference in the outcome or the scope of your errors when using this data, except that use of 1900-1990 data reduces the error rate and better reproduces fig. 1c.
Therefore, the 1900-1990 truncation had no material effect on our analysis. All of our data was also calculated from 1900-present, 1925-present, from 1850-present, from 1800-present, from 1950-present, and across many other time periods, in the hope that we may arrive at a trend that supported your original findings.
Not one of these periods illustrated any plateau in MRAD.
There are several other points that you have misconstrued. The first is that the reasoned criticisms of JMG support your findings, despite his statement that “I am quite convinced by all the problems the authors identified in the analysis performed by Dong et al.”. JMG has provided an insightful comment on our methodology, which we believe results from a simple misunderstanding and has no bearing on the errors in your paper.
The "improperly formed" boxplots of error rates reflect the calculated maximum, median, 95th percentile and 75th percentile of error rates all reaching 100%. That is, the frequency of your zero-rounding errors becomes so high, and error rates below 100% are so rare, there is no longer a 'box' forming in these boxplots.
This was the rationale for including the red and blue lines, showing the overall fraction of data rounded to zero.
As for your comments on the IDL and GRG data, we would highlight several problems. First, you cannot ignore your incorrect pooling of the IDL data because of trends in another database.
There is no evidence that the proposed plateau in MRAD occurs in any single one of these populations. The ‘plateau’ trend that appears when you pool these data is, as we have shown in our code, a result of poor assumptions. You cannot pool four populations in this way, when two of them are only present to 2003 and one to 2006. You have done nothing to address these problems, other than repeating your initial mistake.
Your ‘obvious’ plateau in the GRG is, as we have stated before, based on a single data point. Removal of that data point removes your proposed plateau in these data. It also makes a simple increasing linear model, with no post-1995 plateau in MRAD, the best fit for observed data under the R squared metric, the Bayesian information criteria and the Akaike information criteria.
We find it remarkable that a selected-by-eye regression, with a worse fit than a linear model when Jeanne Calment is removed, is a core supporting argument for your paper.
As we highlighted for JMG, before Jeanne Calment the record-holder for lifespan, Mary Beard held this record. Mary Beard’s record was retracted after being considered valid for 24 years, when it was recognized her extreme age was a straightforward clerical error. As you admit, a similar clerical error for Jeanne Calment would undo your proposal for a significant fall in MRAD.
The GRG acknowledge this inherent unreliability, stating quite clearly that their data are not complete surveys of the oldest old. They also state that many supercentenarian records are subsequently found to be false positives, or are impossible to verify. Therefore, the analysis presented in Nature depends on a single data point, drawn from unreliable data, gathered from incomplete surveys with a high false positive rate, conducted in a small number of countries.
I suggest you ask a statistician whether they consider it good practice to extract species-wide trends from such data.
In contrast, we have based our analysis on surveys of global populations, conducted by national statistical bodies. None of our calculated limits depend on GRG or IDL data.
Your comment that the median MSA we observed, of around 100 years, supports your results is unfounded. That MSA reflects diversity across all countries in the UN database. It better reflects the limit of lifespan in Namibia than in France, or in any of the developed countries you measured. Furthermore, this median is increasing linearly. Like almost all global populations, the MSA is increasing, with no sign of a plateau.
We have provided data on the correspondence between our MSA estimates and observed data: of 311 observations, 310 were consistent with limits calculated under MSA (our figure S2).
Your suggestion that these data are implausible is not based on any apparent analysis. You have not provided any indication why these estimates are incorrect, beyond your own opinion. Where is your code?
We would also highlight that there is nothing historically remarkable about the current length of the Jeanne Calment's lifespan record. We have heard the argument that holding an unbroken lifespan record for two decades somehow suggests an MRAD plateau, an argument you repeated in response to Rozing, Kirkwood and Westendorp (Nature 546, E12 (29 June 2017) doi:10.1038/nature22789).
However, the overall lifespan record was previously held for 17 consecutive years (Mary Kelly, 1964-1981). The male lifespan record was held for 68 consecutive years (Gert Adrians-Boomgaard) and then 21.9 years (Mathew Beard 1985-2007) by a margin of 3 years. None of these broken records indicated a plateau in MRAD.
Neither does the lifespan record of Jeanne Calment. We suggest the Jeanne Calment record simply reflects the uncertain ascertainment, and rarity of adequate documentation, required to validate the oldest-old.
Finally, I consider the findings of Dong et al. are invalid as a result of methodological errors, errors you have not corrected or addressed. Pointing to data you feel supports your view, in this paper or others, does not answer any of the problems raised by ourselves or other authors.
Were our results to entirely support your proposed MRAD plateau this would do nothing to excuse or explain your errors in analysis.
I cannot speak for my (currently traveling) co-author, but I find it disturbing that you have failed to acknowledge or correct even the most basic errors in typesetting and analysis, even those that have little material effect on the result (your Extended Data figure 6 still has incorrect axes, is not the data you said it was in the text, and is still missing all the people who died in May and June).
It seems to many observers that you have committed basic errors that you refuse to acknowledge or repair.
Despite our reproduction of your analysis, you have not provided a single piece of code. We have repeatedly requested this code. Should you ever provide it, in accordance with the editorial guidelines of Nature, I will gladly highlight these errors for you.
In the mean time, I suggest that you reflect on the possibility of honest errors driving your results.
Regards,
Saul Newman
https://f1000researchdata.s3.amazonaws.com/linked/166727.Fig_1_HiRes_2014.tiff