Keywords
autism, pten, macrocephaly, ASD, sensorimotor
autism, pten, macrocephaly, ASD, sensorimotor
The reviewer brought up several good points where we needed to update in the current version. We provided more details on the stimulus that was used in the prepulse inhibition test. We also expanded our discussion to include more comparisons between some of the PTEN knockout models. We also expanded our discussion on some of the caveats in the current study.
See the authors' detailed response to the review by Benjamin K. Yee
See the authors' detailed response to the review by Maarten van den Buuse
See the authors' detailed response to the review by George Leondaritis and Charalampos Brakatselos
Sensorimotor gating is the ability of a sensory stimulus to suppress a motor response1. It can be measured by assessing prepulse inhibition (PPI), wherein a weak auditory stimulus inhibits a startle response that is induced by the following presentation of a loud sound2. Deficits in PPI have been widely reported in various neurological conditions, including autism spectrum disorder (ASD)3–5. Similar to humans, impairments in PPI have been reported in ASD models such as Fmr1 and Cntnap2-knockout (KO) mice; however, the underlying mechanism is unknown6,7. Pten mutant mice are another model of autism and can be used to investigate the connection between a cell signaling pathway commonly implicated in ASD, the PI3K/AKT/mTOR pathway, and specific autistic-like deficits8. Specifically, neuronal subset specific (NS) Pten KO mice have previously been shown to exhibit deficits in repetitive behavior, sociability, and communication, however, prepulse inhibition has not been assessed in this model9,10. In the present study, we used NS-Pten KO mice that exhibit hyperactivation of the PI3K/AKT/mTOR pathway in the cortex, hippocampus, and cerebellum, and assess PPI in order to further elucidate the potential relationship between PI3K/AKT/mTOR signaling and deficits in sensorimotor gating9.
Male and female NS-GFAP-Cre Pten (NS-Pten) heterozygous mice on a FVB based mixed background were obtained from Baylor College of Medicine and have been bred for more than 10 generations at Baylor University. Heterozygous NS-Pten males (n=6) and females (n=12) were used to breed wildtype (WT) and NS-Pten KO pups (RRID: MGI:3714016). The housing for the breeders consisted of two females housed with one male. Genotype was determined from toe clippings taken on postnatal day (PD) 10 (performed by Mouse Genotype, Escondido, CA, USA). On PD 21, animals were weaned and housed with mixed genotype littermates in groups of n=3–5 in cages (Allentown Caging PC7115HT, Allentown, PA, USA) filled with sani-chip bedding (7090 Teklad, Envigo, Somerset, NJ, USA) kept in a room on a 12-hr light/dark diurnal cycle held at 22°C. Mice had ad libitum access to food and water. All animals were tested at 9–10 weeks of age between the hours of 10:00 and 11:30 a.m. Only males were assessed in this study to be in accordance with the literature, as previous behavioral phenotyping studies in the NS-Pten adult model were done in males9,10. Furthermore, a similar study investigating sensorimotor gating in neuron-specific enolase (Nse)-Pten KO mice also only assessed males11. Therefore, in order to make an accurate comparison and to provide similar context with past studies we did not assess females. A total of 29 male mice were assessed, 17 NS-Pten KO and 12 WT mice. The target sample size was determined by, and is in accordance with, the PPI literature11–13. The final sample sizes were as follows: day 1: n=12 WT, n=17 KO, day 2: n=12 WT, n=13 KO, day 3: n=9 WT, n=9 KO. A subset of n= 4 KO mice were excluded from the day 2 analysis and n = 11 mice (3 WT and 8 KO) were excluded from the day 3 analysis due to either a protocol malfunction or the death of the KO animal caused by the occurrence of spontaneous seizures. All test procedures were carried out in compliance with the NIH Guidelines for the Care and Use of Laboratory Animals and were approved by Baylor University’s Institutional Animal Care and Use Committee. Once the experiment concluded, mice were placed into a CO2 chamber and euthanized.
Sensorimotor gating was assessed via the SR-LAB system, which consists of a 15 × 14 × 18 inch isolation cabinet, a plexiglass cylinder (3.2-cm diameter) mounted on a sensor platform, and a speaker that generated white noise, as well as the stimuli (San Diego Instruments, San Diego, CA, USA). The paradigm consisted of three separate testing days: habituation, prepulse inhibition, and startle response, and was conducted as previously described6.
For habituation, the animal was acclimated to the room for 30 minutes then was placed inside the cylinder for a 5-minute habituation period, which was followed by 80 startle stimuli delivered at a fixed interval of 15 seconds. The startle stimulus was a 40 ms, 120 dB white noise burst, with a rise/fall time of less than 1 ms. Prepulse inhibition testing occurred 24 hours after day 1 and consisted of a 5-minute habituation phase that was followed by 20 presentations of a 40 ms, 120 dB white noise burst. In the prepulse phase, mice were presented with 90 trials consisting of three prepulse intensities that were 2, 7, and 12 dB over the 68 dB background noise. The onset of the prepulse occurred 100 ms before the onset of the startle pulse. Each prepulse was 20 ms in duration and were spaced an average of 15 seconds apart (7–23 s). One week after the prepulse session, the startle response was assessed. Following the 5-minute habituation period, the mice were presented with 99 trials of 11 trial types. These included a no stimulus trial and 10 startle stimuli trials ranging from 75–120 dB at 5 dB intervals. The startle stimuli were 40 ms white noise bursts. The 11 trial types were pseudorandomized, with each trial type being presented once in a block of the 11 trials. To eliminate potential confounds during testing, background sound levels were maintained at 68 dB and the experimenter was not present.
GraphPad Prism 7 software (La Jolla, CA) or SPSS 21.0 (IBM, USA) were used to analyze the data. Repeated-measure ANOVAs were run for habituation, prepulse inhibition, and startle response. The within subject factors for habituation were the trials (1–10, 11–20, 21–30, 41–50, 51–60, 61–70, and 71–80) with genotype as the between subjects factor (wildtype and knockout). For prepulse inhibition, the within-subjects factors were the prepulse intensities (2, 7, and 12 dB), with genotype as the between subjects factor. For the startle response, the between-subjects factors were the stimulus intensities (no stimulus, startle at 75, 80, 85, 90, 95, 100, 105, 110, 115, and 120 dB), with the between subject factor of genotype. Due to the skewed data present in the startle response, a log transformation was performed. To provide a general indicator of the overall health of each animal throughout the study, the subject’s weights were assessed with a repeated measures ANOVA that had a within subjects factor of day (weight of the subject for testing days 1, 2, and 3) and a between subjects factor of genotype. No post-hoc tests were performed. A total of n=4 KO mice were excluded from the day 2 analysis and n=11 mice (3 WT and 8 KO) were excluded from the day 3 analysis due to protocol malfunction or death as a result of the severity of the knockout. A value of p < 0.05 was considered significant for each statistical test.
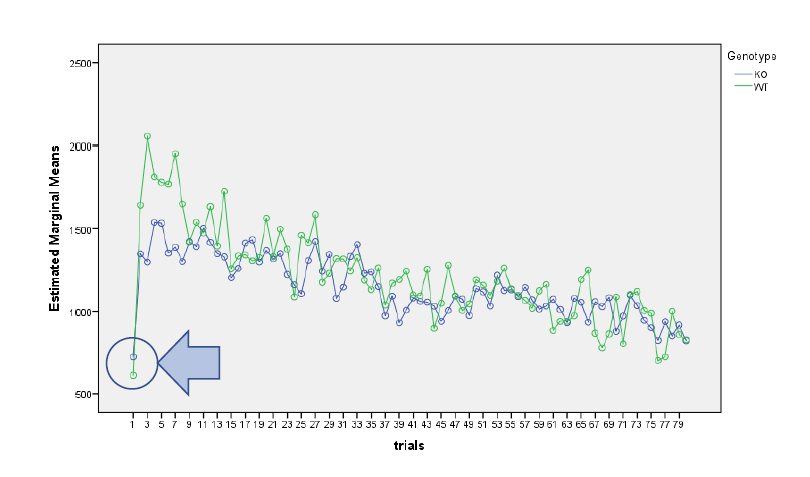
When assessing the sensorimotor gating paradigm, the main effects of genotype was not significant in the ANOVAs of the data obtained on the habituation test on day 1 (F[1,27] = 0.17, p >0.05), the prepulse inhibition test on day 2 (F[1,23] = 2.65, p >0.05) or the startle response test on day 3 (F[1,16] = 2.10, p >0.05). There was also no interaction between genotype and trial for habituation (F[7,189] = 0.91, p >0.05), genotype and prepulse intensity for prepulse inhibition (F[2,46] = 0.71, p >0.05), or genotype and stimulus intensity for the startle response (F[10,160] = .10, p >0.05) (Figure 1a–c). When assessing the weight of each subject throughout the study, no main effect for the within subjects factor of day was found (F[2,30] = .11, p > .05), nor was there a day by genotype interaction (F[2,30] = .17, p > .05). There was also no between subjects effect of genotype (F[1,15] = 1.16, p > .05) (Figure 1d). Raw results for each procedure on each day for every animal are available as Underlying data13.

(a) We found that there was no significant difference in habituation between KO and WT mice (p > 0.05). (b) We found no difference in the percentage of prepulse inhibition between groups following prepulses that were 2, 7, and 12 dB over the 68 dB background noise (p > 0.05). (c) We observed no difference in startle response between NS-Pten KO and WT mice (p > 0.05). (d) We observed no differences in weight between NS-Pten KO and WT mice (p > 0.05). Data are presented as the mean ± standard error of the mean (SEM).
The NS-Pten KO mice did not exhibit significantly different sensorimotor gating from WT mice. A previous study by Kwon et al. (2006) assessed neuron-specific enolase (Nse)-Pten KO mice in a variation of the PPI protocol and reported a decrease in percent inhibition at 4 dB but no differences at 8 or 16 dB11. Our study assessed percent inhibition at 2, 7, and 12 dB, per established protocol, and found no differences at these intensities6. The discrepancy in sensorimotor gating between studies is best attributed to the differences between the Nse and NS-Pten models. For instance, while both Pten models display somatic hypertrophy and have a similar degree of mTOR activation, they display distinct patterns of expression11,14. In the Nse-Pten model, expression is limited to the cortex and hippocampus11. Conversely, in the NS-Pten model, Pten is prominently expressed in the cerebellum in addition to the cortex and hippocampus14. Another primary difference between models is in the timing of the induction of cre expression. Cre expression is induced following neuronal differentiation in Nse-Pten mice, however, in NS-Pten mice cre expression is induced prior to neuronal differentiation11. One outcome of this is a marked deficit in cell migration in NS-Pten mice that is not reported in the Nse-Pten model14,15. Another likely result of the differences in cre expression can be found in the seizure frequency and severity between models. Nse-Pten mice display relatively infrequent short lasting spontaneous seizures11. NS-Pten mice also develop spontaneous seizures however, they are significantly more severe. Indeed, 8 KO animals died in our study due to seizures, despite their age being comparable to Kwon et al.’s (2006) study. Due to the ambiguity surrounding the neurological mechanisms underlying sensorimotor gating, it is difficult to definitively parse out which difference between models best explains our lack of a deficit relative to the literature. Future studies elucidating the mechanisms of PPI would help to address this as would studies that continue to contrast the Nse-Pten and NS-Pten models.
While our study did find compelling evidence that that a premitotic expression of NS-Pten does not result in sensorimotor deficits, there are several potential limitations. For instance, the PPI protocol is inherently stressful. It is possible that anxiety could have masked a sensorimotor gating deficit. Therefore, taking additional steps to minimize animal stress such as increasing the habituation time in the room or further acclimating the animals to the experimental space prior to testing may be beneficial in future studies and ensure that stress does not mask any subtle deficit. However, we have assessed PPI using the same protocol in other strains and found significant differences16. Furthermore, the NS-Pten mice used have previously been shown to have a stress-resistant phenotype9. Therefore, while it is possible that anxiety could have influenced the results, we do not think it is likely. Another potential limitation is the incidence of seizures in the Pten mice. While the seizures are spontaneous and relatively infrequent, they can be severe. It is possible that seizures may have affected the mouse’s behavior in the PPI task, potentially masking a deficit.
Overall, our study provides evidence that the PI3K/AKT/mTOR pathway does not consistently affect sensorimotor gating behaviors in Pten mice. This indicates that any relationship between the two may be contingent on specific developmental stages, expression in particular cell types, or the presence of comorbidities such as seizures. Thus, mTOR appears to have an indirect and nuanced connection to sensorimotor gating processes. This conclusion is supported by a prior study that assessed PPI in a transgenic mouse model of tuberous sclerosis complex, another model of ASD and mTOR hyperactivation, which similarly reported no deficits in prepulse inhibition between WT and KO mice17. Taken together, the literature then indicates that despite mTOR’s contribution to an autistic-like phenotype, it does not unconditionally contribute to the onset of sensorimotor gating deficits, playing a more modest role.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
{kind=link}
Comments on this article Comments (0)