Keywords
microRNA, miRNA, non-coding RNA, enhancer, regulator, macrophage, tuberculosis
This article is included in the Bioinformatics gateway.
This article is included in the Cell & Molecular Biology gateway.
microRNA, miRNA, non-coding RNA, enhancer, regulator, macrophage, tuberculosis
Conventionally, microRNAs (miRNAs) are considered to suppress gene expression through RNA interference (RNAi) by binding complementarily to mRNAs, forming a RISC complex, and causing RNA degradation (reviewed in detail in).1 However, recent studies provide evidence that some miRNAs act in the opposite way – stimulating gene activation. Numerous studies have demonstrated the ability of miRNAs to up-regulate genes by targeting their promoters.2,3 Ago1 from miRNA-Ago complex associates with the Ccnb1 promoter and miR-744 induces enrichment of RNA Pol II and H3K4me3 at the Ccnb1 transcription start site.3 MiRNA let-7i interacts with the TATA-box of IL-2 gene and stimulates transcription initiation by contributing to the assembly of the pre-initiation complex.4 Relatively fewer miRNAs demonstrated the ability to unconventionally target and activate enhancers, thus inducing genes regulated by these enhancers. MiR-24-1 acts as a modulator of the chromatin state of an enhancer. Furthermore, it increases p300 and RNA Pol II binding at the enhancer locus. The miR-24-1 actually originates from the enhancer locus. However, some genes regulated by other enhancers are also expressed at higher levels when miR-24-1 is transfected, and the enhancers of induced genes contain a sequence similar to the seed of the miRNA.5 These observations suggest that other miRNAs might trigger enhancers and activate gene expression.
When miRNAs function as activators in a nucleus, different targeting mechanisms are possible: miRNA:DNA Watson-Crick duplex formation as well as miRNA:DNA Hoogsteen triple helix formation. Nuclear miRNA target prediction tools utilize are based on the idea that miRNA:DNA interaction requires an intact seed region.6 MicroPIR2 predicts targets in mouse and human promoter regions.7 Trident predicts miRNA:DNA Hoogsteen-type base pairings.8 Some tools designed to predict conventional miRNA targets may also be utilized to find nuclear activational targets, e.g. miRanda.9
In this work, we report MIREyA (MIRnas functioning through Enhancer Activation), a pipeline for detection of miRNAs and their gene targets up-regulated through triggering their enhancer in the provided expression dataset. We applied MIREyA in order to identify and characterize activational miRNAs in Mycobacterium tuberculosis (Mtb) infected macrophage dataset. MiRNAs are important regulators in macrophage responses during Mtb infection that may act as host immunity agents as well as a tool exploited by pathogen agents to manipulate host cell pathways.10 Yet, only the classical role of miRNA has been investigated so far in the context of Mtb infection.11–19 Although multiple studies have shown the possibility of the activational role of miRNAs, this potential remains neglected in the study of miRNAs in bacterial infections. MIREyA found several miRNAs, which have not been shown to be functional in TB yet, suggesting it could be useful to find candidate activational miRNAs for further experimental validation. Mir155 has previously been shown to act as a negative regulator of essential mRNAs during TB,20,21 but not as an activator.
MIREyA aims to detect miRNAs with the potential to upregulate a gene via activation of its enhancer. It consists of three major steps:
1) The algorithm detects miRNA bound to the enhancers. This step is implemented with three different approaches described below.
2) The algorithm selects genes regulated by the enhancers selected in step 1). For this step the output from the first step is required as well as a table with enhancer:gene pairs where an enhancer is assumed to regulate the corresponding gene.
3) The algorithm calculates the Spearman’s correlation coefficient (SCC) between the expression levels of miRNAs and genes regulated by corresponding enhancers selected in step 1), and estimates the p-value of the SCC with a Benjamini-Hochberg correction using the number of the miRNA:gene pairs for one miRNA (FDR < 0.05). The input data is gene expression data with sample size ≥ 8 and the output of the previous step.
Figure 1 illustrates the workflow of our algorithm implemented in Python and R.
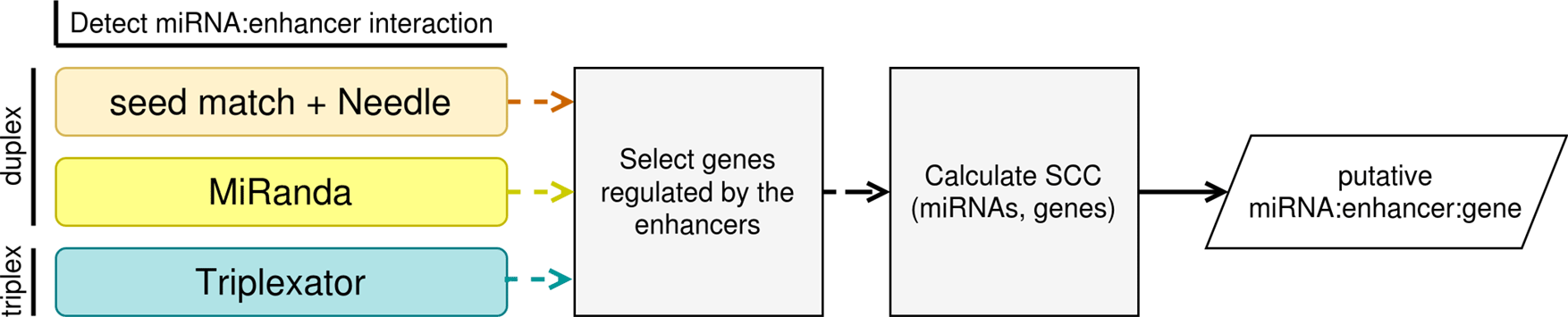
We speculate that in order to activate an enhancer, miRNA should bind to enhancer DNA. Since the mechanism of such binding is unclear, we decided to implement several reasonable prediction strategies. The first two strategies assume that miRNA binds to DNA forming an RNA:DNA double helix, while the third assumes RNA:DNA triple helix formation.
1) The first approach is to select an enhancer containing an exact match of the user-provided seed sequence of a miRNA, then expand each seed by 14 bp of the corresponding mature miRNA and align it to the enhancer with Needle tool22 and keep only enhancers with the percent identity (PI) > 0.5 (PI defined as a percent of matches between miRNA and DNA region).
2) The second approach is to scan miRNA sequences against enhancer sequences and detect potential target sites with MiRanda.9
3) The third approach is to predict RNA:DNA triplexes between miRNAs and enhancers with Triplexator tool.23 We relaxed the error-rate and lower-length-bound Triplexator default parameters in order to adjust the algorithm to work with extremely short miRNA sequences (error-rate=19, lower-length-bound=11).
The approaches are interchangeable, also the user can merge the results of all approaches to reflect multiple mechanisms of potential miRNA:DNA binding.
The main script to run the pipeline is src/run_mireya.py. Input and output files depend on choice of one of previously described approaches to predict miRNA-enhancer interaction and must be specified with the following flags:
-d: approach to predict miRNA-enhancer interaction, accepts one of three possible values: seed_match_needle / miranda / triplexator
-e: path to a fasta file with sequences of enhancers of interest
-o: full path to output directory
-ge: path to .tsv file with gene expression
-me: path to .tsv file with expression of miRNAs of interest
-ei: path to .tsv file with enhancers and corresponding genes they are assumed to regulate
-m: path to a fasta file with sequences of mature miRNAs of interest
Pipeline mode with the first approach to predict miRNA-enhancer interaction (-d seed_match_needle) requires additional input data which must be provided with flags:
-g: path to a folder with fasta files with complete genome of the organism of interest: one chromosome per file
-s: path to a .tsv file with sequences of seeds of miRNAs of interest
-sr: path to .tsv file with reverse complementary sequences of seeds of miRNAs of interest
-eb: path to bed file with coordinates of enhancers of interest
-ms: path to a directory containing directories with mature miRNA sequences: one folder per miRNA containing fasta files with one mature sequence per file. Each directory with mature mirna fastas must be named exactly as in other input files. Names of fasta files will be used further as "mature_mirna" column in result tables.
Examples of command to run the main script, one for each approach to predict miRNA-enhancer interaction (names of files coinside with names of example input files in repository):
1. python src/run_mireya.py -d seed_match_needle -e data/enhancers.macrophages.Mtb.mm9.fasta -o out/seed_match_needle_out/ -ge data/DE_gene_expression.tsv -me data/DE_mirnas_expression.tsv -ei data/enh.gene.assoc.sign.tsv -m data/DE_mirna_mature_seqs.fa -g data/db -s data/seeds_seq_forward_short -sr data/seeds_seq_reverse_compl_short -eb data/enhancers.macrophages.Mtb.bed -ms data/mature_seqs/
2. python src/run_mireya.py -d miranda -e data/enhancers.macrophages.Mtb.mm9.fasta -o out/miranda_out/ -ge data/DE_gene_expression.tsv -me data/DE_mirnas_expression.tsv -ei data/enh.gene.assoc.sign.tsv -m data/DE_mirna_mature_seqs.fa
3. python src/run_mireya.py -d triplexator -e data/enhancers.macrophages.Mtb.mm9.fasta -o out/triplexator_out -ge data/DE_gene_expression.tsv -me data/DE_mirnas_expression.tsv -ei data/enh.gene.assoc.sign.tsv -m data/DE_mirna_mature_seqs.fa
Output file is called mir_enh_gene_trios.tsv. Example output file is placed in the out/ directory of repository.
The pipeline is implemented as Python, R and bash scripts, and can be run with a master script run_mireya.py.
Python>=3.5 and r-base are expected to be pre-installed. Besides, two modes of the pipeline require the following tools installed: MiRanda, Triplexator. The pipeline was tested in Ubuntu and Ubuntu-based linux systems (Ubuntu>=16.04).
We applied MIREyA to three time-series (0, 4, 12, 24, 48, 96 hours) expression datasets (CAGE) of mouse bone marrow-derived macrophages infected with hypervirulent Beijing/W lineage Mycobacterium tuberculosis (Mtb) HN878 strain, 2-3 replicates per time point.24 Each dataset corresponds to the time series after infection for macrophages of different phenotypes: not pre-stimulated (M0), interferon-γ stimulated (M1-polarized) and interleukin-4/interleukin-13 stimulated (M2-polarized). Only differentially expressed (DE) miRNAs and genes were considered. We obtained enhancer-gene interactome from25 where an enhancer is predicted to regulate a gene if their expression levels correlate significantly and they belong to the same topologically associated domain (TAD).
We searched for candidate enhancers targeted by miRNAs with all three methods described previously and merged the results for further steps. CAGE enables one to estimate expression at the promoter level, while enhancers are associated with whole genes. As a proxy of gene expression, we used either an expression value of a promoter with the highest SCC (a) or summed up the expression values of all promoters of the gene (b). To reduce the number of false positive predictions, we selected among miRNA:enhancer duplexes only such cases where (1) a duplex was predicted both by miRanda and Needle-based approach; (2) an enhancer was associated with several genes since in the original paper on miRNA-activated enhancers5 one nuclear miRNA affected expression of multiple genes regulated by the triggered enhancer; (3) the miRNA-gene pair was obtained in both ways (a and b) to estimate expression of the gene. We also added miRNA-gene pairs with highly correlated expression levels (SCC ≥ 0.8) in any of four combinations of two methods to detect miRNA:enhancer interactions (seed match + Needle and miRanda) and two approaches to treat expression of different promoters (a and b). Among predicted miRNA:enhancer triplexes we selected cases where (1) an enhancer was associated with multiple genes; (2) both approaches to estimate gene expression (a and b) yielded this triplex.
We applied MIREyA to three time-series datasets of Mtb-infected macrophages with 3 different phenotypes prior to infection. In M0 macrophages 10 miRNAs were differentially expressed (DE) in at least one time point compared to the state before infection (0h), in M1 there was no DE miRNA, while in M2 only Mir1956 was DE. Figure 2 and Table S1 (Extended data43) represent detected miRNA-enhancer-gene trios for M0 macrophages (Extended data: Table S2 for M2 macrophages43). We investigated the functions of the obtained genes and miRNAs and confirmed that many of them might be involved in the response to Mtb infection. We detected Mir155, which is vastly studied, and known to subvert autophagy in human dendritic cells20 and to be a potential diagnostic marker of active tuberculosis.21 Other miRNAs and their targets which we consider promising for further investigation are summed in Tables 1 and 2. Mir22, Mir221, Mir222 are annotated with high confidence.26 Expression of only some promoters of Klf6 and BC016423 genes correlates with Mir22 expression, but we included them, because they have predicted triplexes with three different enhancers.
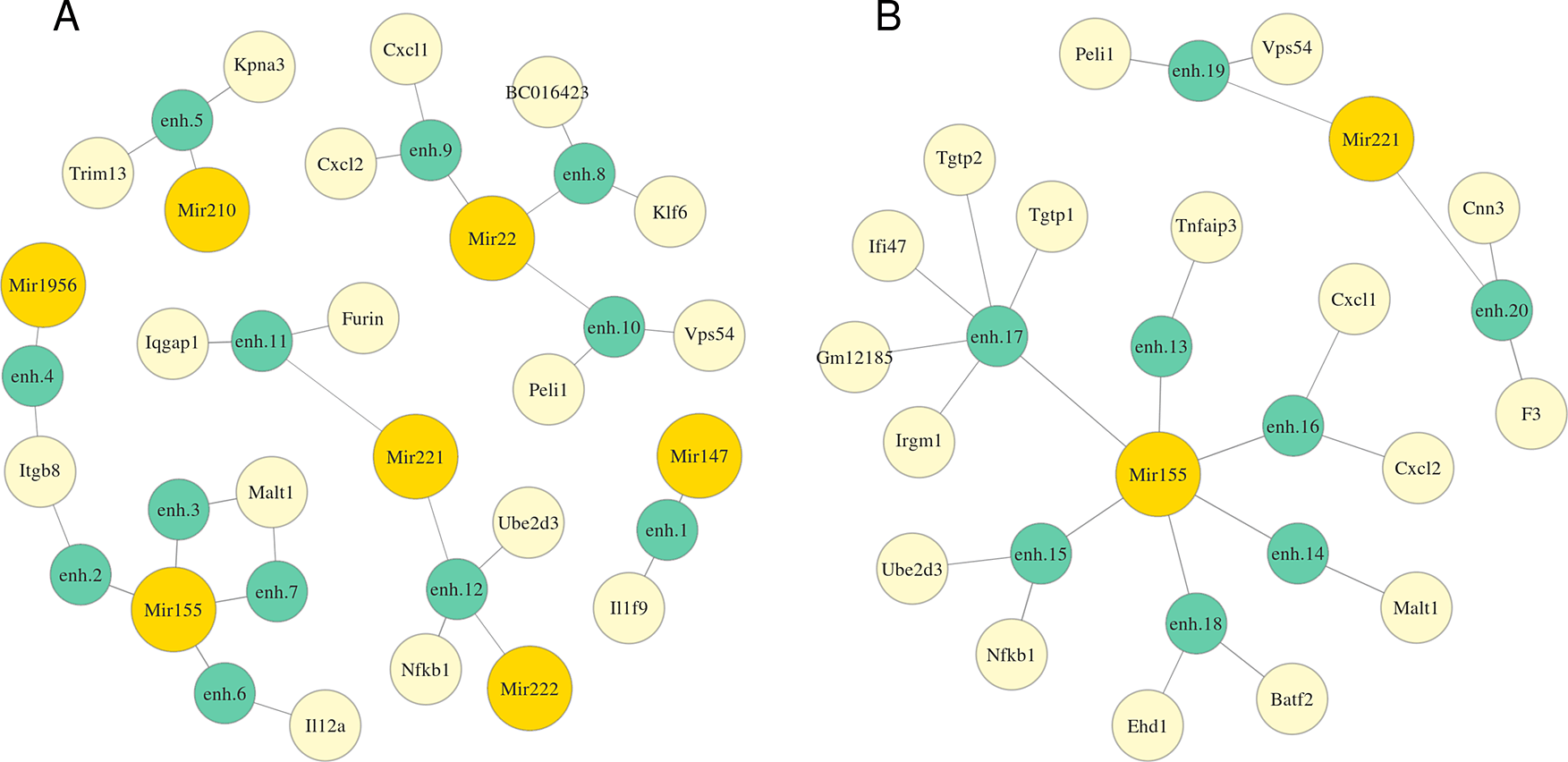
The length of an edge between a miRNA and an enhancer or between an enhancer and a gene is in inverse proportion to the average correlation between the expression of the miRNA and the target gene or the enhancer and the gene respectively. Full coordinates of enhancers are available in Table S1.
| miRNAs | genes | mechanism of interaction of enhancer:miRNA |
|---|---|---|
| Mir1956 | Ccrl2 | duplex |
| Mir1956 | Dotl1 | duplex |
| Mir1956 | Cd14 | duplex |
| Mir1956 | Rab20 | duplex |
| Mir1956 | Ticam1 | duplex |
| Mir1956 | Tnfaip3 (A20) | duplex |
We reconstructed regulatory networks for these miRNAs based on miRNA-enhancer-gene trios and investigated their potential role in the response to Mtb infection. Mir22-activated gene network is highly likely to be involved in Mtb response. Klf6, a potential target of Mir22, is a transcription factor essential for macrophage motility27 and plays an important role in the regulation of macrophage polarization promoting M1 phenotype cooperatively with NF-κB.28 Nfkb1 gene, a putative target of miRNAs Mir221, Mir222 and Mir155, encodes a subunit of NF-κB protein complex, a master transcription factor in macrophage immune responses. The human ortholog of Ube2d3, potentially regulated by the same miRNAs, facilitates polyubiquitination of NFKBIA (a member of the NF-kappa-B inhibitor family) stimulating its subsequent degradation.29 A detected target of both Mir22 and Mir221, Peli1 regulates the NF-κB activity negatively and attenuates the induction of proinflammatory cytokines in T-cells.30 Cxcl1 and Cxcl2, detected targets of both Mir22 and Mir221, are chemoattractants for neutrophils contributing to tissue inflammation.31 Malt1, potentially up-regulated by Mir155, is known to activate NF-κB in lymphocytes.32 Among targets of Mir1956 detected in M2 macrophages dataset we found the Ccrl2 gene encoding a chemokine receptor-like protein which is expressed at high levels in primary neutrophils and primary monocytes. Another Mir1956 target, Dotl1, is an H3K79 methyltransferase which facilitates the expression of IL-6 and IFN-β in macrophages.33 Cd14 leads to NF-κB activation and inflammatory response,34 Cd14 KO mice infected with Mtb are protected due to reduced inflammatory responses at the chronic stage.35 Rab20 plays a role in the maturation and acidification of phagosomes and the fusion of phagosomes with lysosomes during mycobacterial infection.36 Ticam1 is involved in native immunity against pathogens: it interacts specifically with toll-like receptor 3, activates NF-κB.37 Tnfaip3 (A20) is an important regulatory protein that down-regulates NF-κB activity.38
We further investigated protein networks associated with the selected miRNAs. Proteins regulated by miRNAs Mir22, Mir155, Mir221, Mir222 are involved in regulation of NF-κB – a vital orchestrator of the response of the innate immune cells to pathogens39 (Figure 3A). Cxcl1 and Cxcl2 regulated by Mir22 and Mir155 (Figure 3B) are chemokines which signal through CXC receptor 2 to attract neutrophils to the place of inflammation, which is essential to control tissue infection.31
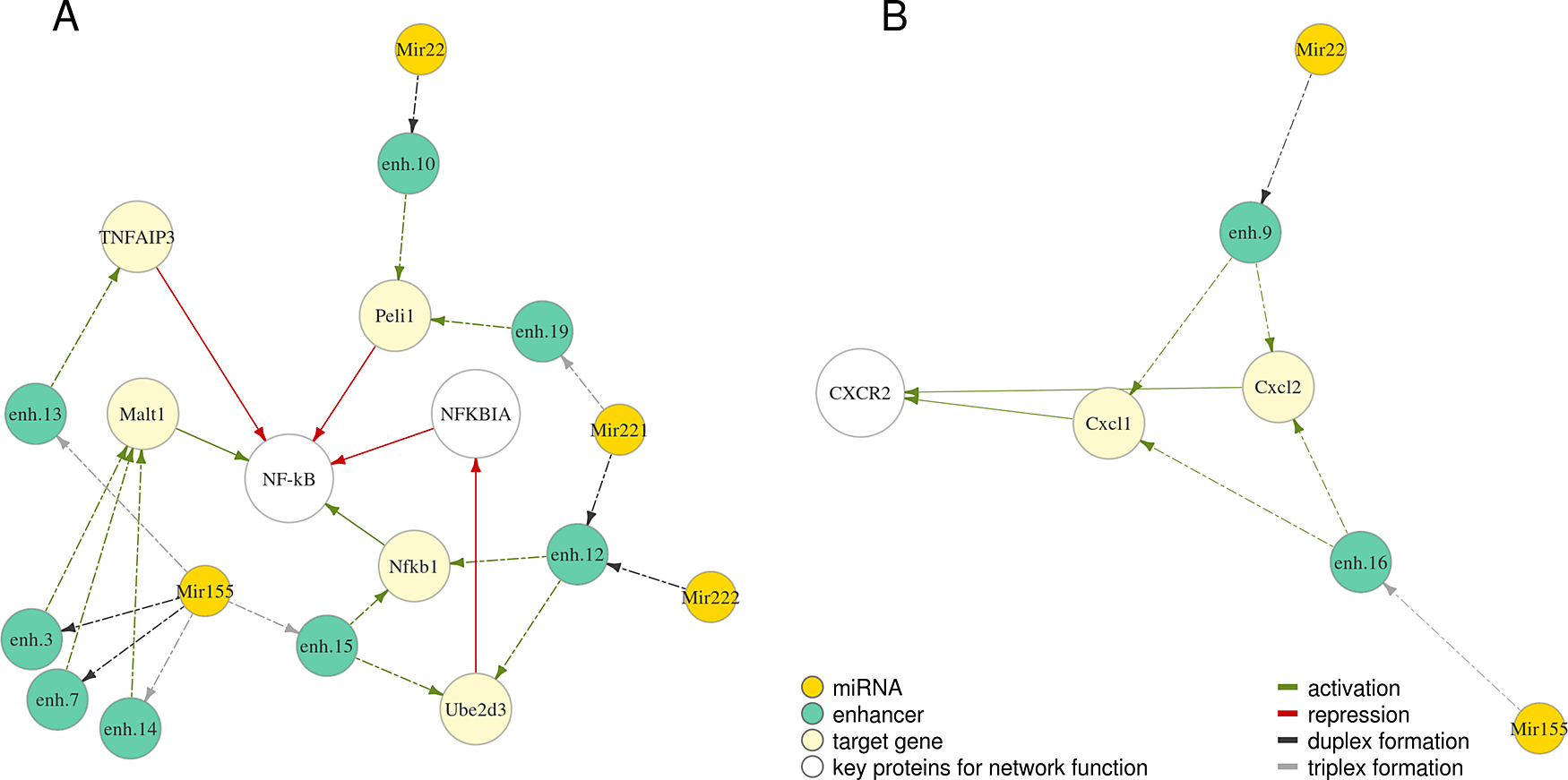
Relationships are depicted with arrows of a different colour: green – activation, red – repression. Dashed arrows indicate predicted relationships, solid – known from published studies and described previously. Full coordinates of enhancers are available in Table S1.
MIREyA aims to easily find candidate activating miRNAs which trigger an enhancer and genes up-regulated by the enhancer. New emerging methods for studying RNA–DNA interactome detect previously unknown miRNAs bound to chromatin40,41 which might be promising for further experimental investigation in terms of understanding complex gene regulation networks in detail. Although high-throughput data on RNA:DNA interactions in several cell types but not macrophages are available now, we could not confirm MiRNAs detected with MIREyA for Mtb infection using either RADICL-seq41 or GRID-seq,42 which is unsurprising since the ncRNA:DNA interactions are highly cell-type-specific.41
Although the implemented algorithm considers multiple aspects of the suggested mechanism of gene activation, several factors remain unaccounted for. To run MIREyA a user requires a priori knowledge of enhancer-gene interactions. A common fast computational approach to determine gene-enhancer pairs is to use genes and enhancers co-localized in the genome, ignoring the long-distance spatial interactions. One of solutions is the approach suggested in:25 to calculate the correlation between expression of genes and enhancers which belong to the same TAD.
Although the exact mechanism of RNA activation of enhancers remains unclear, we do know that miRNA as a mediator facilitates epigenetic modifications in the enhancer region. At this stage, MIREyA does not consider chromatin availability or any other epigenetic information. In order to reduce the number of false positive predictions, nuclear localisation of specific predicted miRNAs should be validated experimentally.
Despite the discussed limitations, using MIREyA we detected several promising miRNA candidates. We suggest that MIREyA provides a promising approach to select miRNAs which up-regulate genes by triggering enhancers for further experimental validation.
Our method extends the study of activational miRNAs and provides a basis for further research. The use case on Mtb-infected macrophages demonstrates the possibility of existence of novel miRNAs up-regulating gene expression.
The CAGE time series expression datasets from the use case are available at https://fantom.gsc.riken.jp/data/ in mm9 Phase 2 release and can be selected to download with FANTOM5 Table Extraction Tool by «macrophage, TB infection» key words.
Zenodo: MIREyA: Extended data, http://doi.org/10.5281/zenodo.4549445.43
This project contains the following extended data:
- Table S1: Detected miRNA-enhancer-gene trios for M0 macrophages
- Table S2: Detected miRNA-enhancer-gene trios for M2 macrophages
Data are available under the terms of the Creative Commons Zero "No rights reserved" data waiver (CC0 1.0 Public domain dedication).
Pipeline available from: https://github.com/veania/MIREyA.
Archived pipeline as at time of publication: https://doi.org/10.5281/zenodo.4635578.44
License: Open Software License 3.0
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)