Buendía E, Marlon M, Parra O et al. Human Proteinase 3, an important autoantigen of c-ANCA associated vasculitis, shares cross-reactive epitopes with serine protease allergens from mites: an in silico analysis [version 2; peer review: 1 approved, 2 approved with reservations]. F1000Research 2022, 10:47 (https://doi.org/10.12688/f1000research.28225.2)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
Human Proteinase 3, an important autoantigen of c-ANCA associated vasculitis, shares cross-reactive epitopes with serine protease allergens from mites: an in silico analysis
[version 2; peer review: 1 approved, 2 approved with reservations]
1Department of Internal Medicine, Universidad del Norte, Barranquilla, Atlantico, 080004, Colombia 2Division of Health Sciences, Universidad del Norte, Barranquilla, Atlantico, 080004, Colombia 3Faculty of Medicine, Universidad de Cartagena, Cartagena, Colombia 4Medical Research group (GINUMED), Universitary Corporation Rafael Núñez, Cartagena, Bolívar, 130001, Colombia 5Department of Internal Medicine, Universidad El Bosque, Bogotá, Cundinamarca, 110111, Colombia 6Departement of Pediatrics, Universidad de Cartagena, Cartagena, Bolívar, 130001, Colombia 7Group of Clinical and Experimental Allergy (GACE), IPS Universitaria, Universidad de Antioquia, Medellín, Antioquia, 050001, Colombia
Background: In autoimmune vasculitis, autoantibodies to Human Proteinase 3 (PR3), a human serine protease, seems to have a role on the inception of c-ANCA associated vasculitis. The origin of this autoreactive response remains unclear. However, for several autoreactive responses, molecular mimicry between environmental antigens and human proteins is key to trigger autoantibodies and finally autoimmunity manifestations. Considering that PR3 is a serine protease and house dust mite (HDM) group 3 allergens share this biochemical activity, the aim of this study was to identify cross-reactive epitopes between serine proteases from human and mites using an in silico approach. Methods: Multi alignment among amino acid sequences of PR3 and HDM group 3 allergens was performed to explore identity and structural homology. ElliPro and BepiPred in silico tools were used to predict B and T cell epitopes. Consurf tool was used to conduct identification of conserved regions in serine proteases family. Results: PR3 and HDM group 3 allergens shared moderate identity and structural homology (root mean square deviation < 1). One B cell cross reactive epitope among serine proteases was identified (29I, 30V, 31G, 32G, 34E, 36K, 37A, 38L, 39A and 54C) and two T cell epitopes. Conclusions: PR3 have structural homology and share cross reactive epitopes with HDM group 3 allergens.
Keywords
Serine proteases, human proteinase 3, house dust mites group 3 allergens, ANCA associated vasculitis, sequence homology, T and B cell epitopes, cross-reactivity, epitope modelling
Corresponding author:
Emiro Buendía
Competing interests:
No competing interests were disclosed.
Grant information:
Buendía, E. received a postdoctoral grant (Fondo Nacional de Financiamiento para la Ciencia, la Tecnología y la Innovación "FRANCISCO JOSÉ DE CALDAS") from Ministerio de Ciencia, Tecnología e Innovación, Colombia (grant number 80740-249-2020).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
The manuscript was revised and refined by a language style corrector. Some sentences in the methods and results section were improved giving more detail and explanations in accord to revisor suggestions.
The manuscript was revised and refined by a language style corrector. Some sentences in the methods and results section were improved giving more detail and explanations in accord to revisor suggestions.
Anti-neutrophil cytoplasmic antibody (ANCA)-associated vasculitis (AAV) is a life-threatening autoimmune disease affecting small vessels, compromising the respiratory mucosa, skin, lung, and the kidney1. This group of small vessel vasculitis includes various diseases: granulomatosis with polyangiitis, microscopic polyangiitis, kidney-limited vasculitis and Eosinophilic granulomatosis with polyangiitis, all of them having in common some degree of autoimmune response to the Human Proteinase 3 protein (PR3). Previous studies have shown that autoantibody binding to PR3 expressed on the neutrophil surface may activate its degranulation, eliciting tissue damage in small vessels and their irrigated organs. Also, while proinflammatory effector T cells have been implicated in vasculitis pathogenesis2, a specific PR3 T cell epitope has not been reported in AAV patients3. PR3 is a serine protease physiologically expressed in human neutrophils. Due to its enzymatic activity, it degrades various intercellular gap-junction proteins and collagen and may play a role in neutrophil transendothelial migration. In addition, this protein is an important autoantigen in AAV, and sera from patients with severe and relapsing forms of the disease can bind it in IgG ELISA assays4–6. Further, although a cause-effect relationship between PR3-autoantibodies and vasculitis is not clearly defined, animal models support a pathogenic role7,8, revealing that they may be involved in disease inception, progression and severity1.
Environmental exposures, specially to microbial components mimicking self-antigens have been proposed as triggers of autoimmunity9,10. Also, in AAV, it has been proposed that an endogenous immune response to a complementary protein to PR3 autoantigen could be implicated in disease inception, and this antisense protein harbors homology to various bacterial peptides11. PR3 crystal structure has been elucidated, and various epitopes are recognized by patients suffering AAV; however, its cross-reactivity with environmental antigens is poorly studied12–14.
Previous studies have shown that specific IgE to some self-proteins have been identified in autoimmune and allergic diseases like lupus, urticaria, dermatitis, allergic pulmonary aspergillosis and have a strong association with disease activity15–18. Some allergens can cross-react with human proteins and participates in autoimmunity inception in pemphigus vulgaris by a “hit-and-run” mechanism, opening the theoretical possibility for a similar mechanism to occur in another autoimmune disease such as AAV19–22.
In the tropics, house dust mites (HDM) are important ubiquitous allergen sources and exposure is perennial, increasing the possibilities of exposure in the general population23, and IgE sensitization to their components24,25. Sensitization to HDM group 3 allergens is common26, as they harbor serine protease activity and conserved structural homology27, making them potential PR3 cross reactive antigens; this has not been explored before. Here, we show in silico data suggesting cross-reactivity and epitope sharing between PR3 and HDM group 3 allergens.
Methods
Searching homologous with BLAST (Basic Local Alignment Search Tool)
The amino acid sequence from the human PR3 (Uniprot accession: P24158) was used as query to perform a search for serine protease homologous reported in allergenic sources: Dermatophagoides pteronyssinus (Der p 3: Accession number P39675), Blomia tropicalis (Blo t 3: A1KXI1), Glycyphagus domesticus (Gly d 3: Q1M2M8), Lepidoglyphus destructor (Led p 3: Q1M2L7) and Tyrophagus putrescentiae (Tyr p 3: C6ZDB5) with the PSI-BLAST tool. Parameters were set as default.
Multiple alignment
Identity among all allergenic sequences homologous to PR3 was analyzed using the Jalview tool2.11.028. First, all allergens and human PR3 codes were used as inputs in the Jalview tool. Second, the T coffee tool was chosen to assess alignment. Third, alignment was displayed as an identity percentage.
Construction of 3D model
The 3D model of Der p 3, a serine protease of Dermatophagoides pteronyssinus was generated by homology in the SWISS-MODEL server using the zymogen catalytic region of human MASP-2 (PDB: 1zjk f) as a template. The 3D model of Der p 3 was loaded into the ProSA-web server29, which was used to analyze its quality.
The model was refined in DeepView v4.1 (energy minimization and rotamer replacements). Its quality was evaluated by several tools, including Ramachandran graphs, WHATIF, QMEAN4 index, and energy values (GROMOS96 force field). For the validation of the Der p 3 structure we used the Minimize Structure option in the UCSF Chimera software, a procedure that adjust the energy and reduce the entropy of the model30.
Three-dimensional structure (PDB: 1FUJ) of the human PR3 serine protease was retrieved from the Protein Data Bank. A cartoon model was created using Pymol software v2.4. Root median square deviation (RMSD) value between Der p 3 and PR3 was calculated using Chimera software v1.030.
B and T cell epitope prediction
ElliPro v3.0 and BepiPred v2.0 tools were used to predict B and T cell epitopes on Der p 331,32. With ElliPro, the 3D structure of Der p 3 was used to predict epitopes. Minimum score and maximum distance (Angstrom) were set to 0.5 and 6, respectively. Epitopes with high conserved rates were visualized in the 3D model. For prediction using BepiPred, an amino acid sequence of Der p 3 was used as input.
Conservation analysis
The 3D structure of Der p 3 was submitted to the ConSurf server to generate evolutionarily related conservation scores to help to identify functional regions in the proteins. HMMER algorithm, 1 iteration, E-value cutoff (0.0001) and UNIREF-90 database was set as default to generate multiple alignment, prior to evolutive analysis. All amino acid sequences in FASTA format were used.
Results
Human PR3 and HDM group 3 allergens exhibited identity and features of the serine protease family
BLAST search identified various serine protease family members from HDM as homologous. The multiple sequence alignment analysis showed that Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3 allergens shared 45% of identity in their aminoacid sequences with PR3. The most conserved region is located between residues 53 to 75, indicating the existence of molecular mimicry (Figure 1). Among the members of HDM group 3 allergens, an identity until 41% was reported (Table 1), and a highly conserved region between residues 40 to 90 was found. When identity between PR3 and each allergen used in study was analyzed, a moderate level of identity was found (30%) (Table 1).
Figure 1. Multiple alignment among PR3 and the HDM allergens belonging to group 3.
An identity of 45% in their amino acid sequences was found.
Table 1. Identity matrix among serine proteases used in study.
All comparisons of PR3 with HDM group 3 allergens showed a moderate identity.
PR3
Der p 3
Blo t 3
Gly d 3
Lep d 3
Tyr p 3
PR3
100
33
27
30
30
27
Der p 3
33
100
48
52
53
43
Blo t 3
27
48
100
58
58
47
Gly d 3
30
52
58
100
99
41
Lep d 3
30
53
58
99
100
41
Tyr p 3
27
43
47
41
41
100
A structural model of Der p 3 was obtained by homology modelling using the 3D structure of PR3 reported in the PDB database. According to modelling, the Der p 3 tertiary structure exhibited a typical fold of serine protease family, conformed by four α-helixes and fifteen β-strands with structural homology with PR3 (RMSD = 0.8) (Figure 2).
Figure 2. Cartoon models of PR3 and Der p 3.
(A) PR3; (B) Der p 3. According to modelling by homology, Der p 3 exhibited a typical fold of serine protease family. (C) Overlapping of 3D model of PR3 and Der p 3. RMSD value of 0.8 is reported, suggesting structural homology.
T and B cell cross-reactive epitopes were predicted between HDM group 3 allergens and PR3
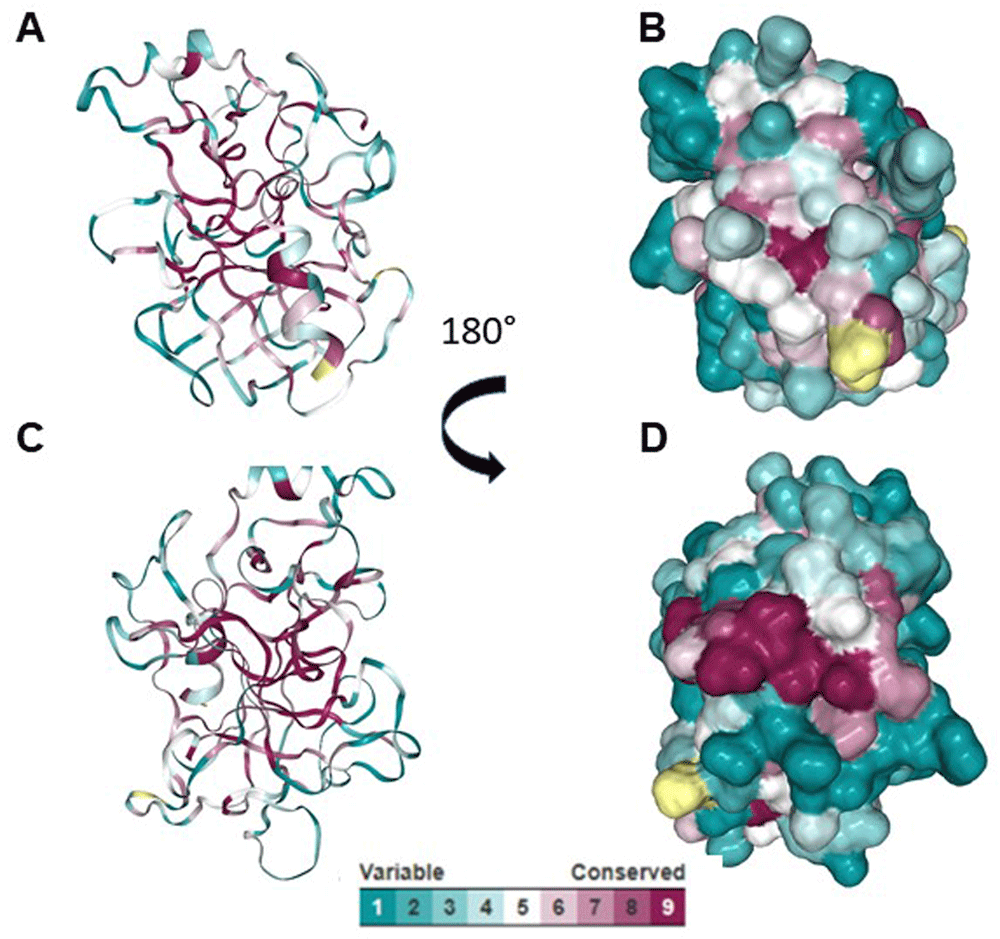
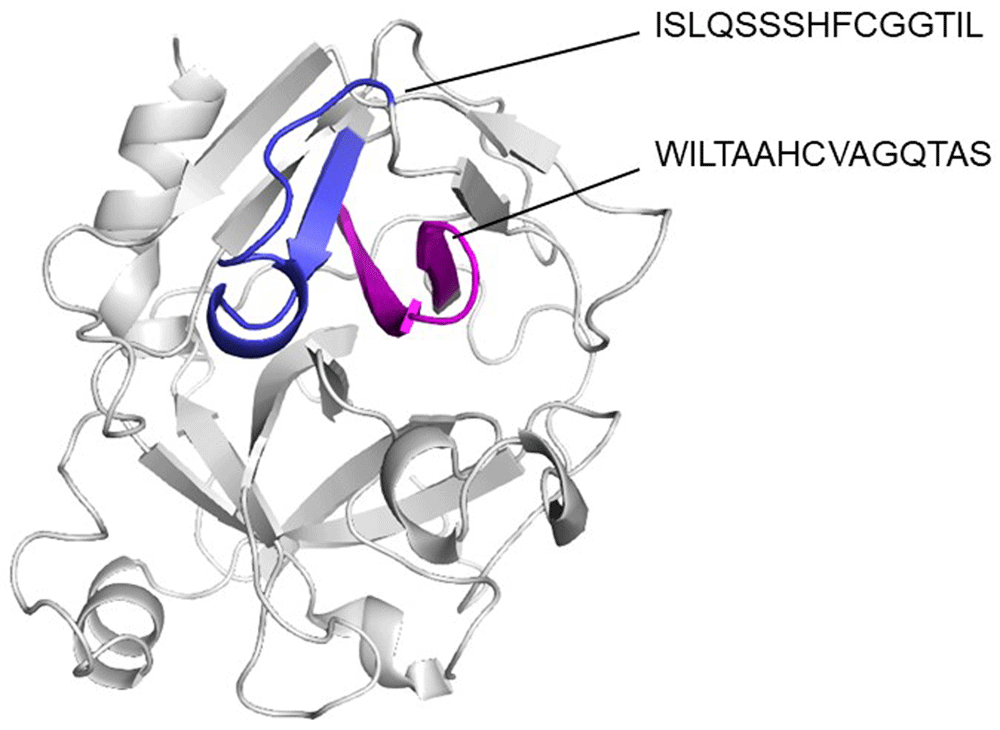
Using ElliPro and BepiPred servers, a cross reactive B cell epitope was predicted on all serine protease used in this study. This epitope is formed by ten residues and is on the N-terminal region, spanning amino acids 29 and 39 with a surface area of 470 Å, not forming part of any domain within the protein. Conservative analysis indicated that the antigenic region predicted was highly conserved in the serine proteases (Figure 3). According to ConSurf analysis, the region covering the cross-reactive epitope is conserved among the serine protease family (Figure 4). T cell epitope prediction identified at least two epitopes with potential cross-reactivity among all sequences analyzed. Both epitopes are located on the first and second β strands: the first epitope spans the 45 to 59 region (ISLQSSSHFCGGTIL); and the second, the 63 to 77 region (WILTAAHCVAGQTAS) (Figure 5; Table 2).
Figure 3.
(A) Surface model of Der p 3, showing area occupied by B cell epitope predicted as cross reactive. (B) Cartoon model showing location of epitope on tridimensional structure. It can be appreciated that the predicted epitope is on a loop spanning residues 29 to 39.
Figure 4. Phylogenetic analysis of the serine proteases using Consurf.
(A and C) Cartoon models showing the conserved region among serine proteases. (B and D) Surface models showing the conserved region among serine proteases.
Figure 5. Cartoon model showing the location of T cell epitopes in their tridimensional structure.
It can be appreciated that predicted epitopes are in a continuous β strands (blue and magenta).
Table 2. T cell epitopes predicted in silico.
Both are conserved among PR3 and HDM group 3 allergens.
Allele
Start
End
Sequence
HLA-DRB1*01:01
45
59
ISLQSSSHFCGGTIL
HLA-DRB1*01:01
63
77
WILTAAHCVAGQTAS
Discussion
In this study we found that PR3 and HDM group 3 serine protease allergens have conserved identity and homology. Also, for the first time, we predicted various T and B cell cross reactive epitopes between them through an in silico approach. PR3 is an important autoantigen in small vessel vasculitis and it seems to participate in disease inception, progression, and severity1. Our results have potential implications for the understanding of autoreactive response in AAV and open the possibility for a new environmental trigger of the autoreactive response in AAV.
In AAV, it has been proposed that autoantibodies directed to a complementary protein to PR3 autoantigen could be implicated in disease inception, and this antisense protein harbors homology to various bacterial peptides11 – a theory named autoantigen complementarity33. However, in epidemiological studies, autoantigen complementarity hypothesis testing has showed conflicting results, since sera from some patients suffering from AAV do not recognize complementary PR3, while others do34–36. Also, molecular mimicry of PR3 protein by infectious microorganism components have been proposed as a possible environmental trigger of the disease based on the reports of infections preceding the manifestations of vasculitis10,37–39, although a cross reactive antigen have not been reported yet.
In their seminal publication, Pendergraft et al. run a BLAST query to find homologues of PR3 protein in microbial or fungal microorganisms, and do not find matching sequences at that time11. However, they do not include Arachnida or other environmental sources of cross-reactivity. In our analysis we find matching PR3 protein sequences with various HDM group 3 serine protease allergens, and at least theoretically this finding could have many implications for the understanding of inception and even diagnosis of autoreactive response in AAV. Recently, Qian et al. have shown that some allergens can cross-react with human proteins19 and participate in autoimmunity inception in pemphigus vulgaris by a “hit-and-run” mechanism22, opening the theoretical possibility for a similar mechanism to occur in another autoimmune disease such as AAV. Similarly, in atopic dermatitis, Valenta and collaborators observe that some patients with severe complications from the disease, had IgE directed to the profilin of the Betula verrucosa, but also to the human homologue40.
In the tropics, HDM are important ubiquitous sources of protease allergens. Exposure is perennial, increasing the possibilities of exposure and IgE sensitization to their components in the general population23–25. Sensitization to HDM group 3 allergens is common26, and they harbor serine protease activity27, a characteristic that make them highly allergenic. Moreover, their conserved structural homology makes them highly immunogenic41,42 and suitable for epitope spreading43. In this context, “hit-and-run” and epitope spreading establish framework mechanisms for environmental allergens with homology to autoantigens to potentially participate in the development of autoimmunity. We speculate that HDM group 3 allergens harbor two characteristics that make them suitable candidates for environmental triggering of AAV: their proteolytic activity that, as other protease allergens, set a tissue damaging microenvironment during antigen recognition41; and molecular homology-epitope sharing with human PR3, that would elicit B cell autoantibody production and autoreactive T cell receptor generation. In conclusion, we observe that PR3 and HDM group 3 serine protease allergens have conserved identity, and for the first time we predict cross-reactive epitopes between them through an in silico approach.
1.
Jennette JC, Falk RJ:
Pathogenesis of antineutrophil cytoplasmic autoantibody-mediated disease.
Nat Rev Rheumatol.
2014; 10(8): 463–73. PubMed Abstract
| Publisher Full Text
2.
Wilde B, Thewissen M, Damoiseaux J, et al.:
T cells in ANCA-associated vasculitis: what can we learn from lesional versus circulating T cells?
Arthritis Res Ther.
2010; 12(1): 204. PubMed Abstract
| Publisher Full Text
| Free Full Text
3.
van der Geld YM, Huitema MG, Franssen CF, et al.:
In Vitro T lymphocyte responses to proteinase 3 (PR3) and linear peptides of PR3 in patients with Wegener's granulomatosis (WG).
Clin Exp Immunol.
2000; 122(3): 504–13. PubMed Abstract
| Publisher Full Text
| Free Full Text
4.
Hogan SL, Falk RJ, Chin H, et al.:
Predictors of relapse and treatment resistance in antineutrophil cytoplasmic antibody-associated small-vessel vasculitis.
Ann Intern Med.
2005; 143(9): 621–31. PubMed Abstract
| Publisher Full Text
5.
Müller A, Voswinkel J, Gottschlich S, et al.:
Human proteinase 3 (PR3) and its binding molecules: implications for inflammatory and PR3-related autoimmune responses.
Ann N Y Acad Sci.
2007; 1109: 84–92. PubMed Abstract
| Publisher Full Text
6.
Specks U:
What you should know about PR3-ANCA. Conformational requirements of proteinase 3 (PR3) for enzymatic activity and recognition by PR3-ANCA.
Arthritis Res.
2000; 2(4): 263–7. PubMed Abstract
| Publisher Full Text
| Free Full Text
7.
Falk RJ, Terrell RS, Charles LA, et al.:
Anti-neutrophil cytoplasmic autoantibodies induce neutrophils to degranulate and produce oxygen radicals In Vitro.
Proc Natl Acad Sci U S A.
1990; 87(11): 4115–9. PubMed Abstract
| Publisher Full Text
| Free Full Text
8.
Xiao H, Heeringa P, Hu P, et al.:
Antineutrophil cytoplasmic autoantibodies specific for myeloperoxidase cause glomerulonephritis and vasculitis in mice.
J Clin Invest.
2002; 110(7): 955–63. PubMed Abstract
| Publisher Full Text
| Free Full Text
10.
Rojas M, Restrepo-Jiménez P, Monsalve DM, et al.:
Molecular mimicry and autoimmunity.
J Autoimmun.
2018; 95: 100–23. PubMed Abstract
| Publisher Full Text
11.
Pendergraft WF 3rd, Preston GA, Shah RR, et al.:
Autoimmunity is triggered by cPR-3(105-201), a protein complementary to human autoantigen proteinase-3.
Nat Med.
2004; 10(1): 72–9. PubMed Abstract
| Publisher Full Text
12.
Fujinaga M, Chernaia MM, Halenbeck R, et al.:
The Crystal Structure of PR3, a Neutrophil Serine Proteinase Antigen of Wegener's Granulomatosis Antibodies.
J Mol Biol.
1996; 261(2): 267–78. PubMed Abstract
| Publisher Full Text
13.
Bruner BF, Vista ES, Wynn DM, et al.:
Anti-neutrophil cytoplasmic antibodies target sequential functional proteinase 3 epitopes in the sera of patients with Wegener’s granulomatosis.
Clin Exp Immunol.
2010; 162(2): 262–70. PubMed Abstract
| Publisher Full Text
| Free Full Text
14.
Kuhl A, Korkmaz B, Utecht B, et al.:
Mapping of conformational epitopes on human proteinase 3, the autoantigen of Wegener's granulomatosis.
J Immunol.
2010; 185(1): 387–99. PubMed Abstract
| Publisher Full Text
15.
Hasni S, Gupta S, Davis M, et al.:
Safety and Tolerability of Omalizumab: A Randomized Clinical Trial of Humanized Anti-IgE Monoclonal Antibody in Systemic Lupus Erythematosus.
Arthritis Rheumatol.
2019; 71(7): 1135–40. PubMed Abstract
| Publisher Full Text
| Free Full Text
16.
Glaser AG, Menz G, Kirsch AI, et al.:
Auto- and cross-reactivity to thioredoxin allergens in allergic bronchopulmonary aspergillosis.
Allergy.
2008; 63(12): 1617–23. PubMed Abstract
| Publisher Full Text
17.
Roesner LM, Heratizadeh A, Wieschowski S, et al.:
α-NAC-Specific Autoreactive CD8+ T Cells in Atopic Dermatitis Are of an Effector Memory Type and Secrete IL-4 and IFN-γ.
J Immunol.
2016; 196(8): 3245–52. PubMed Abstract
| Publisher Full Text
18.
Sánchez J, Sánchez A, Cardona R:
Causal Relationship Between Anti-TPO IgE and Chronic Urticaria by In Vitro and In Vivo Tests.
Allergy Asthma Immunol Res.
2019; 11(1): 29–42. PubMed Abstract
| Publisher Full Text
| Free Full Text
19.
Qian Y, Jeong JS, Maldonado M, et al.:
Cutting Edge: Brazilian pemphigus foliaceus anti-desmoglein 1 autoantibodies cross-react with sand fly salivary LJM11 antigen.
J Immunol.
2012; 189(4): 1535–9. PubMed Abstract
| Publisher Full Text
| Free Full Text
20.
Roesner LM, Ernst M, Chen W, et al.:
Human thioredoxin, a damage-associated molecular pattern and Malassezia-crossreactive autoallergen, modulates immune responses via the C-type lectin receptors Dectin-1 and Dectin-2.
Sci Rep.
2019; 9(1): 11210. PubMed Abstract
| Publisher Full Text
| Free Full Text
21.
Balaji H, Heratizadeh A, Wichmann K, et al.:
Malassezia sympodialis thioredoxin-specific T cells are highly cross-reactive to human thioredoxin in atopic dermatitis.
J Allergy Clin Immunol.
2011; 128(1): 92–9.e4. PubMed Abstract
| Publisher Full Text
22.
Lin L, Moran TP, Peng B, et al.:
Walnut antigens can trigger autoantibody development in patients with pemphigus vulgaris through a "hit-and-run" mechanism.
J Allergy Clin Immunol.
2019; 144(3): 720–8.e4. PubMed Abstract
| Publisher Full Text
| Free Full Text
24.
Zakzuk J, Mercado D, Bornacelly A, et al.:
Hygienic conditions influence sensitization to Blomia tropicalis allergenic components: Results from the FRAAT birth cohort.
Pediatr Allergy Immunol.
2019; 30(2): 172–8. PubMed Abstract
| Publisher Full Text
25.
Acevedo N, Sánchez J, Zakzuk J, et al.:
Particular characteristics of allergic symptoms in tropical environments: follow up to 24 months in the FRAAT birth cohort study.
BMC Pulm Med.
2012; 12: 13. PubMed Abstract
| Publisher Full Text
| Free Full Text
26.
Sánchez-Borges M, Fernandez-Caldas E, Thomas WR, et al.:
International consensus (ICON) on: clinical consequences of mite hypersensitivity, a global problem.
World Allergy Organ J.
2017; 10(1): 14. PubMed Abstract
| Publisher Full Text
| Free Full Text
27.
Stewart GA, Ward LD, Simpson RJ, et al.:
The group III allergen from the house dust mite Dermatophagoides pteronyssinus is a trypsin-like enzyme.
Immunology.
1992; 75(1): 29–35. PubMed Abstract
| Free Full Text
28.
Clamp M, Cuff J, Searle SM, et al.:
The Jalview Java alignment editor.
Bioinformatics.
2004; 20(3): 426–7. PubMed Abstract
| Publisher Full Text
29.
Wiederstein M, Sippl MJ:
ProSA-web: interactive web service for the recognition of errors in three-dimensional structures of proteins.
Nucleic Acids Res.
2007; 35(Web Server issue): W407–10. PubMed Abstract
| Publisher Full Text
| Free Full Text
30.
Pettersen EF, Goddard TD, Huang CC, et al.:
UCSF Chimera--a visualization system for exploratory research and analysis.
J Comput Chem.
2004; 25(13): 1605–12. PubMed Abstract
| Publisher Full Text
31.
Ponomarenko J, Bui HH, Li W, et al.:
ElliPro: a new structure-based tool for the prediction of antibody epitopes.
BMC Bioinformatics.
2008; 9: 514. PubMed Abstract
| Publisher Full Text
| Free Full Text
32.
Jespersen MC, Peters B, Nielsen M, et al.:
BepiPred-2.0: improving sequence-based B-cell epitope prediction using conformational epitopes.
Nucleic Acids Res.
2017; 45(W1): W24–w9. PubMed Abstract
| Publisher Full Text
| Free Full Text
33.
Pendergraft WF 3rd, Pressler BM, Jennette JC, et al.:
Autoantigen complementarity: a new theory implicating complementary proteins as initiators of autoimmune disease.
J Mol Med (Berl).
2005; 83(1): 12–25. PubMed Abstract
| Publisher Full Text
34.
Tadema H, Kallenberg CG, Stegeman CA, et al.:
Reactivity against complementary proteinase-3 is not increased in patients with PR3-ANCA-associated vasculitis.
PLoS One.
2011; 6(3): e17972. PubMed Abstract
| Publisher Full Text
| Free Full Text
35.
Hewins P, Belmonte F, Charles Jennette J, et al.:
Longitudinal studies of patients with ANCA vasculitis demonstrate concurrent reactivity to complementary PR3 protein segments cPR3m and cPR3C and with no reactivity to cPR3N.
Autoimmunity.
2011; 44(2): 98–106. PubMed Abstract
| Publisher Full Text
| Free Full Text
37.
Mezzano S, Valderrama G, Olavarria F, et al.:
Antineutrophil-cytoplasmic-autoantibodies in poststreptococcal nephritis.
Adv Exp Med Biol.
1993; 336: 449–53. PubMed Abstract
| Publisher Full Text
38.
Hellmich B, Ehren M, Lindstaedt M, et al.:
Anti-MPO-ANCA-positive microscopic polyangiitis following subacute bacterial endocarditis.
Clin Rheumatol.
2001; 20(6): 441–3. PubMed Abstract
| Publisher Full Text
39.
Pudifin DJ, Duursma J, Gathiram V, et al.:
Invasive amoebiasis is associated with the development of anti-neutrophil cytoplasmic antibody.
Clin Exp Immunol.
1994; 97(1): 48–51. PubMed Abstract
| Publisher Full Text
| Free Full Text
40.
Valenta R, Duchêne M, Pettenburger K, et al.:
Identification of profilin as a novel pollen allergen; IgE autoreactivity in sensitized individuals.
Science.
1991; 253(5019): 557–60. PubMed Abstract
| Publisher Full Text
41.
Cayrol C, Duval A, Schmitt P, et al.:
Environmental allergens induce allergic inflammation through proteolytic maturation of IL-33.
Nat Immunol.
2018; 19(4): 375–85. PubMed Abstract
| Publisher Full Text
42.
Westernberg L, Schulten V, Greenbaum JA, et al.:
T-cell epitope conservation across allergen species is a major determinant of immunogenicity.
J Allergy Clin Immunol.
2016; 138(2): 571–8.e7. PubMed Abstract
| Publisher Full Text
| Free Full Text
43.
Burastero SE:
T-cell receptor-mediated cross-reactivity to different allergens is driven by recognition of homologous, phylogenetically conserved epitopes.
J Allergy Clin Immunol.
2016; 138(4): 1237. PubMed Abstract
| Publisher Full Text
1
Department of Internal Medicine, Universidad del Norte, Barranquilla, Atlantico, 080004, Colombia 2
Division of Health Sciences, Universidad del Norte, Barranquilla, Atlantico, 080004, Colombia 3
Faculty of Medicine, Universidad de Cartagena, Cartagena, Colombia 4
Medical Research group (GINUMED), Universitary Corporation Rafael Núñez, Cartagena, Bolívar, 130001, Colombia 5
Department of Internal Medicine, Universidad El Bosque, Bogotá, Cundinamarca, 110111, Colombia 6
Departement of Pediatrics, Universidad de Cartagena, Cartagena, Bolívar, 130001, Colombia 7
Group of Clinical and Experimental Allergy (GACE), IPS Universitaria, Universidad de Antioquia, Medellín, Antioquia, 050001, Colombia
Buendía, E. received a postdoctoral grant (Fondo Nacional de Financiamiento para la Ciencia, la Tecnología y la Innovación "FRANCISCO JOSÉ DE CALDAS") from Ministerio de Ciencia, Tecnología e Innovación, Colombia (grant number 80740-249-2020).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Buendía E, Marlon M, Parra O et al. Human Proteinase 3, an important autoantigen of c-ANCA associated vasculitis, shares cross-reactive epitopes with serine protease allergens from mites: an in silico analysis [version 2; peer review: 1 approved, 2 approved with reservations]. F1000Research 2022, 10:47 (https://doi.org/10.12688/f1000research.28225.2)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
Tanaka H. Reviewer Report For: Human Proteinase 3, an important autoantigen of c-ANCA associated vasculitis, shares cross-reactive epitopes with serine protease allergens from mites: an in silico analysis [version 2; peer review: 1 approved, 2 approved with reservations]. F1000Research 2022, 10:47 (https://doi.org/10.5256/f1000research.81402.r136303)
To elucidate the origin of the autoreactive response of human proteinase 3 (PR3), the authors examined molecular similarity between environmental antigens and PR3. They found that some house dust mites showed structural homology to PR3. Some environmental antigens, such as
... Continue reading
To elucidate the origin of the autoreactive response of human proteinase 3 (PR3), the authors examined molecular similarity between environmental antigens and PR3. They found that some house dust mites showed structural homology to PR3. Some environmental antigens, such as house dust mites, may be attributable to the production of PR3-ANCA.
This is an interesting paper. The results obtained is informative enough for readers.
It is nice to add some comments or speculation about how the results obtained are relevant in real-world clinical practice, particularly the etiology and treatment of patients with C-ANCA-related vasculitis. This issue is very relevant for physicians who treat patients with ANCA-related vasculitis.
Is the work clearly and accurately presented and does it cite the current literature?
Yes
Is the study design appropriate and is the work technically sound?
Yes
Are sufficient details of methods and analysis provided to allow replication by others?
Yes
If applicable, is the statistical analysis and its interpretation appropriate?
Not applicable
Are all the source data underlying the results available to ensure full reproducibility?
Yes
Are the conclusions drawn adequately supported by the results?
Yes
Competing Interests: No competing interests were disclosed.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Tanaka H. Reviewer Report For: Human Proteinase 3, an important autoantigen of c-ANCA associated vasculitis, shares cross-reactive epitopes with serine protease allergens from mites: an in silico analysis [version 2; peer review: 1 approved, 2 approved with reservations]. F1000Research 2022, 10:47 (https://doi.org/10.5256/f1000research.81402.r136303)
Erban T. Reviewer Report For: Human Proteinase 3, an important autoantigen of c-ANCA associated vasculitis, shares cross-reactive epitopes with serine protease allergens from mites: an in silico analysis [version 2; peer review: 1 approved, 2 approved with reservations]. F1000Research 2022, 10:47 (https://doi.org/10.5256/f1000research.81402.r124319)
Buendia et al. investigated in their manuscript possible link between human proteinase 3 (PR3) and house dust mite allergen protease that is denoted as allergen group 3. Authors suggest that similarity between PR3 and the group 3 mite allergens may
... Continue reading
Buendia et al. investigated in their manuscript possible link between human proteinase 3 (PR3) and house dust mite allergen protease that is denoted as allergen group 3. Authors suggest that similarity between PR3 and the group 3 mite allergens may be link to autoimmune vasculitis. Methodology and conclusions are based on structural homology based on in silico analysis. I believe that recommendations will improve the manuscript, which is of interest.
Because methodology of the study is based on in silico tools, it is necessary to consider that the function is “possible” and needs to be confirmed in future.
Please note, that group 3 mite allergens become subject of debate in the domestic mites, that include both hose dust (HDMs) and stored product mite (SPMs) species. Immortally, authors should clarify that allergens of HDMs and SPMs were included in the study. In addition, these allergens are present in mite feces and, therefore, they are abundant in the environment. Thus, introduction should be improved.
It was found that occurrence of trypsin allergens in mites differs in species and strains. The expression of group 3 mite allergens was observed to be affected by presence of intracellular symbionts in mites. There is suggested link to PAR2 (PAR-2) in mites (– for instance, see: https://doi.org/10.1016/j.jprot.2021.1043561). The affected pathway in mites can be affected also in humans. Could be a suggested link to PR3 in this regard? e.g. “immune functions of PR3 with respect to PR3 modulation of cell activation via cleavage of protease-activated receptor-2 (PAR-2)” - Ann N Y Acad Sci. 2007 Aug;1109:84-92. doi: 10.1196/annals.1398.0102.
Authors can also consider the suggested “Orchestration of an uncommon maturation cascade of the house dust mite protease allergen quartet” (https://doi.org/10.3389/fimmu.2014.001383)
Comment to approach
Authors used different sequences of group 3 mite allergens to predict similarity. It should be clear whether “pro” or mature form was used in analyses. It is recommended that authors will consider change “identity” to similarity. Usually, high identity is “only” to partial sequence. It is important to consider overall similarity.
In initial stage, authors used multiple Grp3 sequences, i.e.: Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3. Is it possible to report similarity?
In further analysis, authors used Der p 3 sequence. Incidentally, research showed that D. pteronyssinus is depleted of this allergen in mite feces, although it is high-abundance in D. farinae – for instance, see J Proteomics. 2017 Jun 6;162:11-19 doi: 10.1016/j.jprot.2017.04.0214
Is the work clearly and accurately presented and does it cite the current literature?
Partly
Is the study design appropriate and is the work technically sound?
Partly
Are sufficient details of methods and analysis provided to allow replication by others?
Yes
If applicable, is the statistical analysis and its interpretation appropriate?
Partly
Are all the source data underlying the results available to ensure full reproducibility?
Yes
Are the conclusions drawn adequately supported by the results?
Partly
References
1. Erban T, Klimov P, Harant K, Talacko P, et al.: Label-free proteomic analysis reveals differentially expressed Wolbachia proteins in Tyrophagus putrescentiae: Mite allergens and markers reflecting population-related proteome differences. Journal of Proteomics. 2021; 249. Publisher Full Text 2. Müller A, Voswinkel J, Gottschlich S, Csernok E: Human proteinase 3 (PR3) and its binding molecules: implications for inflammatory and PR3-related autoimmune responses.Ann N Y Acad Sci. 2007; 1109: 84-92 PubMed Abstract | Publisher Full Text 3. Dumez ME, Herman J, Campizi V, Galleni M, et al.: Orchestration of an uncommon maturation cascade of the house dust mite protease allergen quartet.Front Immunol. 2014; 5: 138 PubMed Abstract | Publisher Full Text 4. Erban T, Harant K, Hubert J: Detailed two-dimensional gel proteomic mapping of the feces of the house dust mite Dermatophagoides pteronyssinus and comparison with D. farinae : Reduced trypsin protease content in D. pteronyssinus and different isoforms. Journal of Proteomics. 2017; 162: 11-19 Publisher Full Text
Competing Interests: No competing interests were disclosed.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Erban T. Reviewer Report For: Human Proteinase 3, an important autoantigen of c-ANCA associated vasculitis, shares cross-reactive epitopes with serine protease allergens from mites: an in silico analysis [version 2; peer review: 1 approved, 2 approved with reservations]. F1000Research 2022, 10:47 (https://doi.org/10.5256/f1000research.81402.r124319)
Selvaraj C. Reviewer Report For: Human Proteinase 3, an important autoantigen of c-ANCA associated vasculitis, shares cross-reactive epitopes with serine protease allergens from mites: an in silico analysis [version 2; peer review: 1 approved, 2 approved with reservations]. F1000Research 2022, 10:47 (https://doi.org/10.5256/f1000research.31220.r90330)
NOTE: it is important to ensure the information in square brackets after the title is included in this citation.
Reviewer Report09 Aug 2021
Chandrabose Selvaraj,
Department of Bioinformatics, Computer Aided Drug Design and Molecular Modeling Lab, Alagappa University, Karaikudi, Tamilnadu, 630003, India
Primary check on this manuscript for language refinement is mandatory as I see that it is not satisfactory, and moreover, there are more typographical errors. I suggest the authors go with language
... Continue reading
Reviewer Recommendation: Major Revision
Primary check on this manuscript for language refinement is mandatory as I see that it is not satisfactory, and moreover, there are more typographical errors. I suggest the authors go with language and grammar checks.
The following sentence seems incomplete and difficult to understand, reframe it “Here, allergens such as: Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3 shared an identity of 45% among them according to multiple sequence alignment"
Why have authors selected PR-3 for the homologous evaluation study? Since the 3-D structure is available in the PDB database?
What basis authors have selected allergenic sources such as Dermatophagoides pteronyssinus (Der p 3: Accession number P39675), Blomia tropicalis (Blo t 3: A1KXI1), Glycyphagus domesticus (Gly d 3: Q1M2M8), Lepidoglyphus destructor (Led p 3: Q1M2L7) and Tyrophagus putrescentiae (Tyr p 3:C6ZDB5) in this study? In BLAST it did not show the above mentioned allergens.
Did authors find the sequence similarity analysis of target sequence Der p 3 with its template; if so provide that information more in detail.
Swiss-Modeler does not show the 1FUJ as template in template searching, and then what basis authors have checked RMSD calculation between the modeled structure and IFUJ as template?
In methodology part authors have mentioned that the modeled structure was subjected to validation, but lack results and interpretation. Provide more in detail.
Provide abbreviation for AAV
Is the residues present in the epitope region responsible for the antigenic effect?
In epitope prediction, the author only provides the sequence and length of the epitopes, if the author provides the information on score value, identity and whether it is present in any domain or motif it could be more informative.
Is the work clearly and accurately presented and does it cite the current literature?
Yes
Is the study design appropriate and is the work technically sound?
Partly
Are sufficient details of methods and analysis provided to allow replication by others?
No
If applicable, is the statistical analysis and its interpretation appropriate?
No
Are all the source data underlying the results available to ensure full reproducibility?
Partly
Are the conclusions drawn adequately supported by the results?
Partly
Competing Interests: No competing interests were disclosed.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Selvaraj C. Reviewer Report For: Human Proteinase 3, an important autoantigen of c-ANCA associated vasculitis, shares cross-reactive epitopes with serine protease allergens from mites: an in silico analysis [version 2; peer review: 1 approved, 2 approved with reservations]. F1000Research 2022, 10:47 (https://doi.org/10.5256/f1000research.31220.r90330)
Emiro Buendía, Department of Internal Medicine, Universidad del Norte, Barranquilla, 080004, Colombia
03 Feb 2022
Author Response
Emiro Buendía, Division of Health Sciences, Universidad del Norte, Barranquilla, Colombia.
Dear reviewer, thanks for the reading of the paper, we have taken note of all your comments, questions
...
Continue readingEmiro Buendía, Division of Health Sciences, Universidad del Norte, Barranquilla, Colombia.
Dear reviewer, thanks for the reading of the paper, we have taken note of all your comments, questions and suggestions. Our responses are bellow:
1. Reviewer: Primary check on this manuscript for language refinement is mandatory as I see that it is not satisfactory, and moreover, there are more typographical errors. I suggest the authors go with language and grammar checks.
Answer: We appreciate your suggestion. The manuscript was revised and refined by a language style corrector.
2. Reviewer: The following sentence seems incomplete and difficult to understand, reframe it “Here, allergens such as: Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3 shared an identity of 45% among them according to multiple sequence alignment"
Answer: We appreciate your suggestion. A new sentence was added: “Multiple sequence alignment showed that Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3 allergens shared 45% of identity in their aminoacid sequences with PR3. The most conserved region is located between residues 53 to 75, indicating the existence of molecular mimicry.”
3. Reviewer: Why have authors selected PR-3 for the homologous evaluation study? Since the 3-D structure is available in the PDB database?
Answer: We appreciate your question. We selected PR3 for the study because it is the most important autoantigen in ANCA + vasculitis. Since, this human protein is a serine protease, in the sequence alignment analysis we seek for other serine proteases in mites, because it is a previously described environmentally ubiquitous serine protease source. In our study we selected the 3D model of PR3 from the Protein Data Bank as mentioned in your observation because its structure was already deciphered, the structural homology analysis was done keeping in mind that its serine protease folding could make more possible for us to model homologous serine in mites.
4. Reviewer: What basis authors have selected allergenic sources such as Dermatophagoides pteronyssinus (Der p 3: Accession number P39675), Blomia tropicalis (Blo t 3: A1KXI1), Glycyphagus domesticus (Gly d 3: Q1M2M8), Lepidoglyphus destructor (Led p 3: Q1M2L7) and Tyrophagus putrescentiae (Tyr p 3:C6ZDB5) in this study? In BLAST it did not show the above-mentioned allergens.
Answer: We appreciate your question. These allergens were chosen on the basis that they belong to the serine protease family, and our hypothesis was that the sensitization to these allergens is cross-reactive with the PR3 autoantigen. So, search was limited using terms: American house dust mite (taxid:6954). In this way, results were only about mites, an important allergenic source.
5. Reviewer: Did authors find the sequence similarity analysis of target sequence Der p 3 with its template; if so provide that information more in detail.
Answer: We appreciate your question. Yes, the binary alignment showed 33% identity between the template amino acid sequences, and Der p 3. An image of the alignment was uploaded in the new manuscript version.
6. Reviewer: Swiss-Modeler does not show the 1FUJ as template in template searching, and then what basis authors have checked RMSD calculation between the modeled structure and IFUJ as template?
Answer: We appreciate your question. For modelling the mite allergen Der p 3 structure, we used the zymogen catalytic region of MASP-2, a lectin binding mannose from human (PDB: 1zjk f), as a template because 1FUJ code was not listed in Swiss-Modeler query results for this protein. Next we calculated the RMSD based on the predicted structure of the mite Der p3 protease and the previously resolved structure of the human PR3. The following sentence was improved to answer your question in the method section of the paper (Construction of 3D model): “The 3D model of Der p 3, a serine protease of Dermatophagoides pteronyssinus was generated by homology in the SWISS-MODEL server using the zymogen catalytic region of human MASP-2 (PDB: 1zjk f) as a template. The 3D model of Der p 3 was loaded into the ProSA-web server 29, which was used to analyze its quality”.
7. Reviewer: In methodology part authors have mentioned that the modeled structure was subjected to validation, but lack results and interpretation. Provide more in detail.
Answer: We appreciate your suggestion. We added the next sentence in the method section (Construction of 3D model): For the validation of the Der p 3 structure we used the Minimize Structure option in the UCSF Chimera software, a procedure that adjust the energy (entropy) of the model.
8. Reviewer: Provide abbreviation for AAV.
Answer: We appreciate your suggestion. ANCA Associated Vasculitis (AAV) was included in the corrected paper text.
9. Reviewer: Is the residues present in the epitope region responsible for the antigenic effect?
Answer: We appreciate your question. We think that the residues found in the epitope region are involved in the cross reactivity between the studied serine protease and hypothesized have a role in autoimmunity inception. But this are preliminary results awaiting experimental validation.
10. Reviewer: In epitope prediction, the author only provides the sequence and length of the epitopes, if the author provides the information on score value, identity and whether it is present in any domain or motif it could be more informative.
Answer: We appreciate your suggestion. The B epitope is part of the N-terminal region of PR3 and is not within any domain of the protein. The T epitope spanned between residues 45-59 is located on the first and second beta lamina. While the second epitope T (63-77) is located in the third beta sheet, and first alpha helix in the protein structure. In the result section (T and B cell cross-reactive epitopes were predicted between HDM group 3 allergens and PR3), two sentences were improved to consider your suggestion.
Emiro Buendía, Division of Health Sciences, Universidad del Norte, Barranquilla, Colombia.
Dear reviewer, thanks for the reading of the paper, we have taken note of all your comments, questions and suggestions. Our responses are bellow:
1. Reviewer: Primary check on this manuscript for language refinement is mandatory as I see that it is not satisfactory, and moreover, there are more typographical errors. I suggest the authors go with language and grammar checks.
Answer: We appreciate your suggestion. The manuscript was revised and refined by a language style corrector.
2. Reviewer: The following sentence seems incomplete and difficult to understand, reframe it “Here, allergens such as: Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3 shared an identity of 45% among them according to multiple sequence alignment"
Answer: We appreciate your suggestion. A new sentence was added: “Multiple sequence alignment showed that Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3 allergens shared 45% of identity in their aminoacid sequences with PR3. The most conserved region is located between residues 53 to 75, indicating the existence of molecular mimicry.”
3. Reviewer: Why have authors selected PR-3 for the homologous evaluation study? Since the 3-D structure is available in the PDB database?
Answer: We appreciate your question. We selected PR3 for the study because it is the most important autoantigen in ANCA + vasculitis. Since, this human protein is a serine protease, in the sequence alignment analysis we seek for other serine proteases in mites, because it is a previously described environmentally ubiquitous serine protease source. In our study we selected the 3D model of PR3 from the Protein Data Bank as mentioned in your observation because its structure was already deciphered, the structural homology analysis was done keeping in mind that its serine protease folding could make more possible for us to model homologous serine in mites.
4. Reviewer: What basis authors have selected allergenic sources such as Dermatophagoides pteronyssinus (Der p 3: Accession number P39675), Blomia tropicalis (Blo t 3: A1KXI1), Glycyphagus domesticus (Gly d 3: Q1M2M8), Lepidoglyphus destructor (Led p 3: Q1M2L7) and Tyrophagus putrescentiae (Tyr p 3:C6ZDB5) in this study? In BLAST it did not show the above-mentioned allergens.
Answer: We appreciate your question. These allergens were chosen on the basis that they belong to the serine protease family, and our hypothesis was that the sensitization to these allergens is cross-reactive with the PR3 autoantigen. So, search was limited using terms: American house dust mite (taxid:6954). In this way, results were only about mites, an important allergenic source.
5. Reviewer: Did authors find the sequence similarity analysis of target sequence Der p 3 with its template; if so provide that information more in detail.
Answer: We appreciate your question. Yes, the binary alignment showed 33% identity between the template amino acid sequences, and Der p 3. An image of the alignment was uploaded in the new manuscript version.
6. Reviewer: Swiss-Modeler does not show the 1FUJ as template in template searching, and then what basis authors have checked RMSD calculation between the modeled structure and IFUJ as template?
Answer: We appreciate your question. For modelling the mite allergen Der p 3 structure, we used the zymogen catalytic region of MASP-2, a lectin binding mannose from human (PDB: 1zjk f), as a template because 1FUJ code was not listed in Swiss-Modeler query results for this protein. Next we calculated the RMSD based on the predicted structure of the mite Der p3 protease and the previously resolved structure of the human PR3. The following sentence was improved to answer your question in the method section of the paper (Construction of 3D model): “The 3D model of Der p 3, a serine protease of Dermatophagoides pteronyssinus was generated by homology in the SWISS-MODEL server using the zymogen catalytic region of human MASP-2 (PDB: 1zjk f) as a template. The 3D model of Der p 3 was loaded into the ProSA-web server 29, which was used to analyze its quality”.
7. Reviewer: In methodology part authors have mentioned that the modeled structure was subjected to validation, but lack results and interpretation. Provide more in detail.
Answer: We appreciate your suggestion. We added the next sentence in the method section (Construction of 3D model): For the validation of the Der p 3 structure we used the Minimize Structure option in the UCSF Chimera software, a procedure that adjust the energy (entropy) of the model.
8. Reviewer: Provide abbreviation for AAV.
Answer: We appreciate your suggestion. ANCA Associated Vasculitis (AAV) was included in the corrected paper text.
9. Reviewer: Is the residues present in the epitope region responsible for the antigenic effect?
Answer: We appreciate your question. We think that the residues found in the epitope region are involved in the cross reactivity between the studied serine protease and hypothesized have a role in autoimmunity inception. But this are preliminary results awaiting experimental validation.
10. Reviewer: In epitope prediction, the author only provides the sequence and length of the epitopes, if the author provides the information on score value, identity and whether it is present in any domain or motif it could be more informative.
Answer: We appreciate your suggestion. The B epitope is part of the N-terminal region of PR3 and is not within any domain of the protein. The T epitope spanned between residues 45-59 is located on the first and second beta lamina. While the second epitope T (63-77) is located in the third beta sheet, and first alpha helix in the protein structure. In the result section (T and B cell cross-reactive epitopes were predicted between HDM group 3 allergens and PR3), two sentences were improved to consider your suggestion.
Competing Interests:No competing interests were disclosed.Close
Emiro Buendía, Department of Internal Medicine, Universidad del Norte, Barranquilla, 080004, Colombia
03 Feb 2022
Author Response
Emiro Buendía, Division of Health Sciences, Universidad del Norte, Barranquilla, Colombia.
Dear reviewer, thanks for the reading of the paper, we have taken note of all your comments, questions
...
Continue readingEmiro Buendía, Division of Health Sciences, Universidad del Norte, Barranquilla, Colombia.
Dear reviewer, thanks for the reading of the paper, we have taken note of all your comments, questions and suggestions. Our responses are bellow:
1. Reviewer: Primary check on this manuscript for language refinement is mandatory as I see that it is not satisfactory, and moreover, there are more typographical errors. I suggest the authors go with language and grammar checks.
Answer: We appreciate your suggestion. The manuscript was revised and refined by a language style corrector.
2. Reviewer: The following sentence seems incomplete and difficult to understand, reframe it “Here, allergens such as: Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3 shared an identity of 45% among them according to multiple sequence alignment"
Answer: We appreciate your suggestion. A new sentence was added: “Multiple sequence alignment showed that Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3 allergens shared 45% of identity in their aminoacid sequences with PR3. The most conserved region is located between residues 53 to 75, indicating the existence of molecular mimicry.”
3. Reviewer: Why have authors selected PR-3 for the homologous evaluation study? Since the 3-D structure is available in the PDB database?
Answer: We appreciate your question. We selected PR3 for the study because it is the most important autoantigen in ANCA + vasculitis. Since, this human protein is a serine protease, in the sequence alignment analysis we seek for other serine proteases in mites, because it is a previously described environmentally ubiquitous serine protease source. In our study we selected the 3D model of PR3 from the Protein Data Bank as mentioned in your observation because its structure was already deciphered, the structural homology analysis was done keeping in mind that its serine protease folding could make more possible for us to model homologous serine in mites.
4. Reviewer: What basis authors have selected allergenic sources such as Dermatophagoides pteronyssinus (Der p 3: Accession number P39675), Blomia tropicalis (Blo t 3: A1KXI1), Glycyphagus domesticus (Gly d 3: Q1M2M8), Lepidoglyphus destructor (Led p 3: Q1M2L7) and Tyrophagus putrescentiae (Tyr p 3:C6ZDB5) in this study? In BLAST it did not show the above-mentioned allergens.
Answer: We appreciate your question. These allergens were chosen on the basis that they belong to the serine protease family, and our hypothesis was that the sensitization to these allergens is cross-reactive with the PR3 autoantigen. So, search was limited using terms: American house dust mite (taxid:6954). In this way, results were only about mites, an important allergenic source.
5. Reviewer: Did authors find the sequence similarity analysis of target sequence Der p 3 with its template; if so provide that information more in detail.
Answer: We appreciate your question. Yes, the binary alignment showed 33% identity between the template amino acid sequences, and Der p 3. An image of the alignment was uploaded in the new manuscript version.
6. Reviewer: Swiss-Modeler does not show the 1FUJ as template in template searching, and then what basis authors have checked RMSD calculation between the modeled structure and IFUJ as template?
Answer: We appreciate your question. For modelling the mite allergen Der p 3 structure, we used the zymogen catalytic region of MASP-2, a lectin binding mannose from human (PDB: 1zjk f), as a template because 1FUJ code was not listed in Swiss-Modeler query results for this protein. Next we calculated the RMSD based on the predicted structure of the mite Der p3 protease and the previously resolved structure of the human PR3. The following sentence was improved to answer your question in the method section of the paper (Construction of 3D model): “The 3D model of Der p 3, a serine protease of Dermatophagoides pteronyssinus was generated by homology in the SWISS-MODEL server using the zymogen catalytic region of human MASP-2 (PDB: 1zjk f) as a template. The 3D model of Der p 3 was loaded into the ProSA-web server 29, which was used to analyze its quality”.
7. Reviewer: In methodology part authors have mentioned that the modeled structure was subjected to validation, but lack results and interpretation. Provide more in detail.
Answer: We appreciate your suggestion. We added the next sentence in the method section (Construction of 3D model): For the validation of the Der p 3 structure we used the Minimize Structure option in the UCSF Chimera software, a procedure that adjust the energy (entropy) of the model.
8. Reviewer: Provide abbreviation for AAV.
Answer: We appreciate your suggestion. ANCA Associated Vasculitis (AAV) was included in the corrected paper text.
9. Reviewer: Is the residues present in the epitope region responsible for the antigenic effect?
Answer: We appreciate your question. We think that the residues found in the epitope region are involved in the cross reactivity between the studied serine protease and hypothesized have a role in autoimmunity inception. But this are preliminary results awaiting experimental validation.
10. Reviewer: In epitope prediction, the author only provides the sequence and length of the epitopes, if the author provides the information on score value, identity and whether it is present in any domain or motif it could be more informative.
Answer: We appreciate your suggestion. The B epitope is part of the N-terminal region of PR3 and is not within any domain of the protein. The T epitope spanned between residues 45-59 is located on the first and second beta lamina. While the second epitope T (63-77) is located in the third beta sheet, and first alpha helix in the protein structure. In the result section (T and B cell cross-reactive epitopes were predicted between HDM group 3 allergens and PR3), two sentences were improved to consider your suggestion.
Emiro Buendía, Division of Health Sciences, Universidad del Norte, Barranquilla, Colombia.
Dear reviewer, thanks for the reading of the paper, we have taken note of all your comments, questions and suggestions. Our responses are bellow:
1. Reviewer: Primary check on this manuscript for language refinement is mandatory as I see that it is not satisfactory, and moreover, there are more typographical errors. I suggest the authors go with language and grammar checks.
Answer: We appreciate your suggestion. The manuscript was revised and refined by a language style corrector.
2. Reviewer: The following sentence seems incomplete and difficult to understand, reframe it “Here, allergens such as: Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3 shared an identity of 45% among them according to multiple sequence alignment"
Answer: We appreciate your suggestion. A new sentence was added: “Multiple sequence alignment showed that Der p 3, Blo t 3, Gly d 3, Led p 3 and Tyr p 3 allergens shared 45% of identity in their aminoacid sequences with PR3. The most conserved region is located between residues 53 to 75, indicating the existence of molecular mimicry.”
3. Reviewer: Why have authors selected PR-3 for the homologous evaluation study? Since the 3-D structure is available in the PDB database?
Answer: We appreciate your question. We selected PR3 for the study because it is the most important autoantigen in ANCA + vasculitis. Since, this human protein is a serine protease, in the sequence alignment analysis we seek for other serine proteases in mites, because it is a previously described environmentally ubiquitous serine protease source. In our study we selected the 3D model of PR3 from the Protein Data Bank as mentioned in your observation because its structure was already deciphered, the structural homology analysis was done keeping in mind that its serine protease folding could make more possible for us to model homologous serine in mites.
4. Reviewer: What basis authors have selected allergenic sources such as Dermatophagoides pteronyssinus (Der p 3: Accession number P39675), Blomia tropicalis (Blo t 3: A1KXI1), Glycyphagus domesticus (Gly d 3: Q1M2M8), Lepidoglyphus destructor (Led p 3: Q1M2L7) and Tyrophagus putrescentiae (Tyr p 3:C6ZDB5) in this study? In BLAST it did not show the above-mentioned allergens.
Answer: We appreciate your question. These allergens were chosen on the basis that they belong to the serine protease family, and our hypothesis was that the sensitization to these allergens is cross-reactive with the PR3 autoantigen. So, search was limited using terms: American house dust mite (taxid:6954). In this way, results were only about mites, an important allergenic source.
5. Reviewer: Did authors find the sequence similarity analysis of target sequence Der p 3 with its template; if so provide that information more in detail.
Answer: We appreciate your question. Yes, the binary alignment showed 33% identity between the template amino acid sequences, and Der p 3. An image of the alignment was uploaded in the new manuscript version.
6. Reviewer: Swiss-Modeler does not show the 1FUJ as template in template searching, and then what basis authors have checked RMSD calculation between the modeled structure and IFUJ as template?
Answer: We appreciate your question. For modelling the mite allergen Der p 3 structure, we used the zymogen catalytic region of MASP-2, a lectin binding mannose from human (PDB: 1zjk f), as a template because 1FUJ code was not listed in Swiss-Modeler query results for this protein. Next we calculated the RMSD based on the predicted structure of the mite Der p3 protease and the previously resolved structure of the human PR3. The following sentence was improved to answer your question in the method section of the paper (Construction of 3D model): “The 3D model of Der p 3, a serine protease of Dermatophagoides pteronyssinus was generated by homology in the SWISS-MODEL server using the zymogen catalytic region of human MASP-2 (PDB: 1zjk f) as a template. The 3D model of Der p 3 was loaded into the ProSA-web server 29, which was used to analyze its quality”.
7. Reviewer: In methodology part authors have mentioned that the modeled structure was subjected to validation, but lack results and interpretation. Provide more in detail.
Answer: We appreciate your suggestion. We added the next sentence in the method section (Construction of 3D model): For the validation of the Der p 3 structure we used the Minimize Structure option in the UCSF Chimera software, a procedure that adjust the energy (entropy) of the model.
8. Reviewer: Provide abbreviation for AAV.
Answer: We appreciate your suggestion. ANCA Associated Vasculitis (AAV) was included in the corrected paper text.
9. Reviewer: Is the residues present in the epitope region responsible for the antigenic effect?
Answer: We appreciate your question. We think that the residues found in the epitope region are involved in the cross reactivity between the studied serine protease and hypothesized have a role in autoimmunity inception. But this are preliminary results awaiting experimental validation.
10. Reviewer: In epitope prediction, the author only provides the sequence and length of the epitopes, if the author provides the information on score value, identity and whether it is present in any domain or motif it could be more informative.
Answer: We appreciate your suggestion. The B epitope is part of the N-terminal region of PR3 and is not within any domain of the protein. The T epitope spanned between residues 45-59 is located on the first and second beta lamina. While the second epitope T (63-77) is located in the third beta sheet, and first alpha helix in the protein structure. In the result section (T and B cell cross-reactive epitopes were predicted between HDM group 3 allergens and PR3), two sentences were improved to consider your suggestion.
Competing Interests:No competing interests were disclosed.Close
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles



Comments on this article Comments (0)