Keywords
drug resistance, Plasmodium falciparum, antifolate, artemisinin, genetic markers, Eritrea
This article is included in the Pathogens gateway.
drug resistance, Plasmodium falciparum, antifolate, artemisinin, genetic markers, Eritrea
A statement on the in vivo findings which resulted in treatment policy change was added in the introduction.
The reference number under the ethical statement was changed.
See the authors' detailed response to the review by Sam L Nsobya
See the authors' detailed response to the review by Olusola Ojurongbe
Malaria is a major vector-borne disease, endemic in 87 tropical and sub-tropical countries, causing over 400,000 deaths yearly (WHO World Malaria Report 2020). Eritrea, which is situated in the Horn of Africa, has experienced a significant decline in deaths and cases of malaria over the past 20 years (WHO World Malaria Report 2019). This reduction, according to the Ministry of Health (MOH) reports, is mainly due to extensive interventions employed towards the control of malaria since the establishment of the Eritrea National Malaria Control Program (NMCP) in 1995.1 Working hand-in-hand with Roll Back Malaria (RBM) collaborators and stakeholders,2 NMCP set up a combination of strategies including integrated vector management (IVM), early diagnosis and prompt treatment3 consequently leading to a remarkable decrease in incidence and mortality rates, following the gruesome 1998 malaria epidemic in the country.4 The disease is generally endemic in the Western lowlands of Gash Barka, Anseba, Debub and Semenawi Keih Bahri (Northern Red Sea) zobas (regions) whereas the Central highlands and Eastern lowlands of Maekel and Debubawi Keih Bahri (Southern Red Sea) zobas respectively have unstable, seasonal transmission. July–September is the common rainy season and hence malaria transmission peaks between October–November in a majority of the endemic areas while in the Coastal region the rainy season mostly occurs between December–January leading to a heightened transmission in March–April.5,6 Malaria transmission in the western lowlands is highly seasonal, peaking during the rainy season (June – November), and declining considerably during the dry season (December – June).7,8 The risk of malaria infection is estimated at 70 infective bites per year in the western lowlands, with high entomologic inoculation rates during the rainy season and little transmission during the dry season.7 Generally, malaria prevalence in the western lowlands is highly focal, with a low parasitemia proportion of 1.9% (ranging from 0.4% to 3.8%).9 Additionally, the prevalence of malaria infection cases in Eritrea occurs across all age groups, unlike a majority of Sub-Saharan Africa where malaria occurs mainly in children below five years and pregnant women.10 About three-quarters of confirmed malaria cases in Eritrea are caused by Plasmodium falciparum and the remaining one-quarter is attributed to Plasmodium vivax, as well as small proportions of mixed infections (WHO World Malaria Report 2014). Currently, case management in Eritrea exclusively entails World Health Organisation (WHO) recommended first line treatment of uncomplicated malaria using artesunate-amodiaquine (AS-AQ), an artemisinin-based combination therapy (ACT) adopted in 2007, while quinine (Q) has been used for severe cases of infection since 2002 (WHO World Malaria Report 2014). Monitoring for drug resistance plays a major role in governing the efficacy of antimalarials, which subsequently influences their use in a population.
The emergence of drug resistance, especially among P. falciparum parasites, is a major hindrance to malaria control due to its increasing prevalence to essentially all antimalarials including sulfadoxine-pyrimethamine (SP) and lately artemisinins (ARTs).11 Genetic markers are invaluable tools in screening and detection of drug resistance, in addition to predicting the efficacy of antimalarials.12 Sulfadoxine-pyrimethamine P. falciparum resistance (SPR), which is well-studied, results from the occurrence and accumulation of mutations in the dihydrofolate reductase gene (Pfdhfr) and in the dihydropteorate synthase gene (Pfdhps) leading to a gradual reduction of sensitivity to pyrimethamine and sulfadoxine respectively.13 In vitro and in vivo studies have shown that SPR is mainly associated with point mutations at codons N51I, C59R, S108N and I164L of Pfdhfr and S436A, A437G, K540E, A581G and A613S of Pfdhps.14,15 Various combinations of these mutations have been used to classify SP resistant parasites according to different levels of resistance i.e. partially-, fully- or super resistant parasites and this has subsequently affected SP treatment policy. Partial resistance is demonstrated by a combination of triple mutant Pfdhfr, N51I, C59R, S108N and Pfdhps, A437G whereas full resistance is shown by a combination of triple mutant Pfdhfr, N51I, C59R, S108N and double mutant Pfdhfr, A437G, K540E. Finally, the sextuple mutant genotype involving a combination of triple mutant Pfdhfr, N51I, C59R, S108N and triple mutant Pdhfr, A437G, K540E and A581G defines super resistance.16
The development of artemisinin (ART) resistant P. falciparum parasites was first independently described in Western Cambodia, South East Asia.17 To date, resistance is commonly associated with five non-synonymous mutations including M476I, Y493H, R539T, I543T, and C580Y in the propeller domain of P. falciparum kelch 13 gene (Pfk13).18,19 ART resistance is primarily characterized by delayed parasite clearance rates in clinical studies as well as reduced in vitro drug susceptibility of the ring stage of parasite development.20,21 Additionally, there is the likely existence of a large reservoir of Pfk13-mutations globally, evidenced by the presence of non-synonymous mutations not associated with delayed parasite, especially in SSA. This has been demonstrated in a previous survey that involved screening of over 1000 African P. falciparum infections across various sites.22 Considering the significant malaria control interventions accomplished in Eritrea, this pilot study aimed at availing supplementary molecular data by screening for SP and ART resistance-associated mutations from a cohort of patients, treated with first line AS-AQ, visiting selected hospitals located in malaria endemic regions of Eritrea. Generally, despite WHO’s change in treatment policy from the chloroquine (CQ) - sulfadoxine-pyrimethamine (SP) combination, adopted in 2002 to ACT in 2007, due to 50% in vivo treatment failure rate23 little is documented on the point mutations underlying SPR Pfdhfr and Pfdhps using genetic markers. Therefore, one objective of this study was to genotype point mutations associated with SP resistance, since SP was previously used for malaria case management in the general population of Eritrea. Unlike most malaria-endemic countries of Sub-Saharan Africa, widespread SP implementation as an intermittent preventive treatment for children (IPTc) or pregnancy (IPTp), has not been done in Eritrea (WHO World Malaria Report 2020). Another objective was to genotype point mutations associated with ART resistance in the PfK-13 genetic marker, as well as, other emerging non-synonymous (nsy) mutations not associated with ART resistance, reported in previous studies. A continuous detection for ART-resistance using genetic markers is important to keep track of changes at the genetic level.
The ethical approval for this study was obtained from the Eritrea Institute of Technology, Research and Postgraduate Studies (RPS) Ethics Review Committee (Reference no. RPS/169/14) and the Ethics Review Board of the National Commission for Higher Education, Eritrea (NCHE) (Reference no. BHEAIL/3/656-658/14).
This study was conducted at three hospitals located in the western malaria-endemic lowlands of Eritrea: Adi Quala Hospital, Adi Quala (14°38′07′′N, 38°50′03′′E) in Zoba Debub, Keren Hospital, Keren (15°46′40′′N, 38°27′03′′E) in Zoba Anseba and Gash Barka Referral Hospital, Barentu (15°06′20′′N, 37°35′26′′E) in Zoba Gash Barka. Three time ranges were employed for the study at the three hospitals: from 1st July to 31st August 2014 for Adi Qualla Hospital, 16th July to 15th September 2014 for Keren Hospital and 15th August to 1st October 2014 for Gash Barka Referral Hospital.
All patients aged above twenty (20) years, with fever of temperatures > 37.5°C at the consultation visit or a history of fever within the previous 24 hours, were confirmed for malaria infection by microscopic examination of 10% Giemsa-stained thin and thick blood slides. After written assent was given, patients with P. falciparum mono-infection of initial density between 1000 and 100,000 asexual parasites per microliter (uL) of blood were included in the study. From these patients, those who also tested positive on rapid diagnostic testing of P. falciparum histidine-rich protein 2 (CareStart® Pf/Pv, Access Bio, USA) were included. Other inclusion criteria of the study were: absence of an antimalarial treatment history in the previous two weeks, availability for follow-up after treatment prescription, absence of clinical and parasitological evidence of complicated malaria, absence of confirmed pregnancy or breastfeeding, and absence of a history of allergy or adverse reactions to the administered antimalarials or concomitant illnesses.
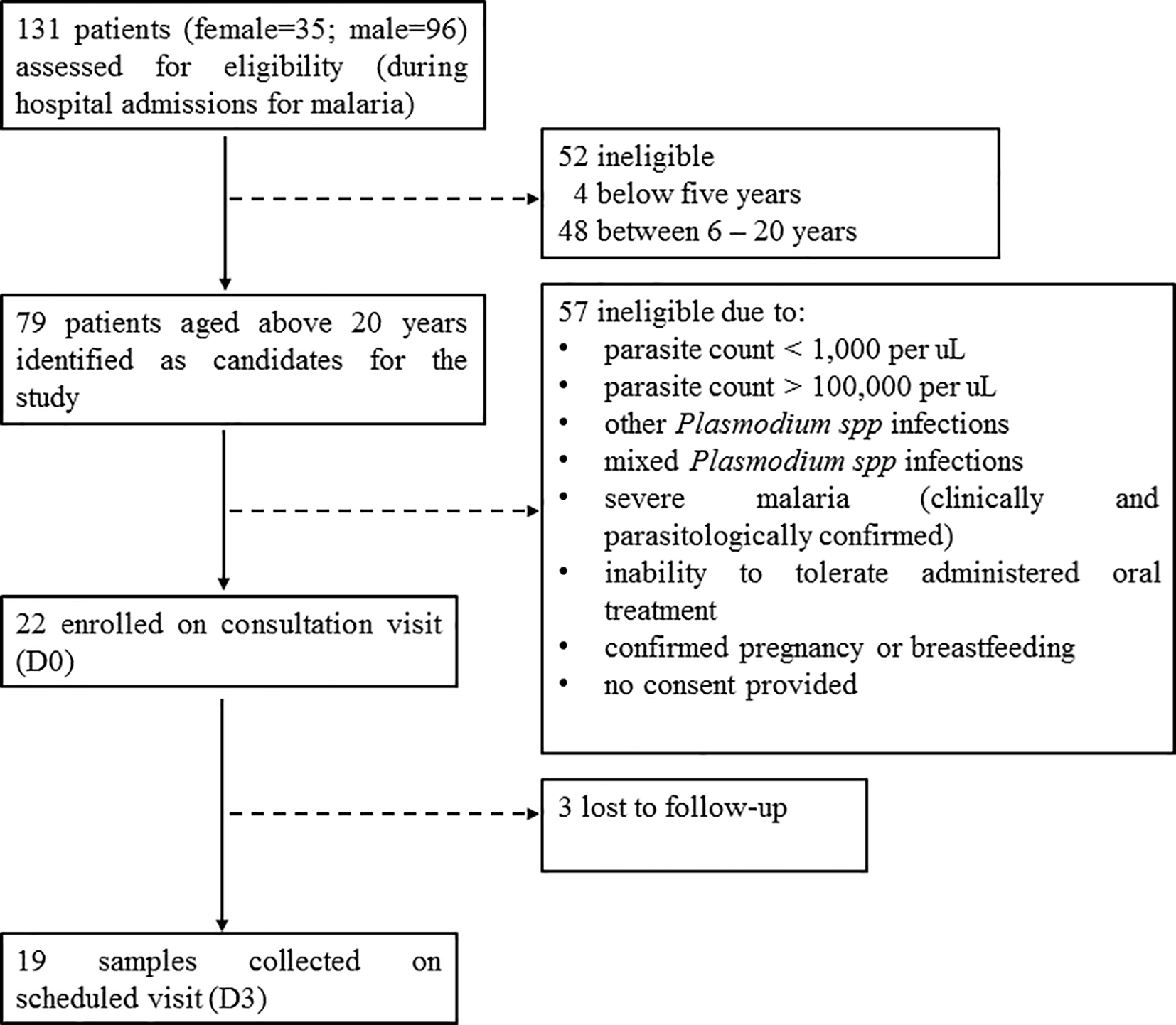
A total of 131 patients (female=35; male=96) with slide-confirmed malaria infection were admitted at the three out-patient hospital sites during the study period.24 From these, 79 patients aged above 20 years (female=23; male=56), were identified as candidates for the study.24 However, 22 patients (female=8; male=14) who met the other inclusion criteria were enrolled in the study on the consultation visit (Day 0) (Figure 1). For all enrolled patients, microscopic examination and rapid diagnostic testing of P. falciparum mono-infection was conducted on the consultation visit and on the subsequent scheduled visits. Oral tablets of AS-AQ were prescribed once a day for three consecutive days, according to the treatment regimen. Treatment administration after Day 0 (D0) was unsupervised on D1 and D2, and sample collection was scheduled on D3. On the scheduled visit (D3), 19 blood samples (AQH=10 samples; KH=3 samples; GBH=6 samples) were successfully collected, while three patients (female=2; male=1) were lost to follow-up (Figure 1).
The blood samples collected were spotted on Whatman 903TM paper (GE Healthcare Bioscience Corp.), stored in individual plastic bags with silica desiccant and transported for further molecular studies at the Institute for Biotechnology Research (IBR) in Jomo Kenyatta University of Agriculture and Technology (JKUAT), Kenya, for genomic DNA extraction and PCR amplification. On each filter paper, the date of sample collection, and the patients’ serial numbers depicted as the hospital code preceding a unique identifier (i.e. AQHxxx, KHxxx, GBHxxx), were recorded.
Genomic DNA extraction was performed on the dried blood spot (DBS) samples using Schneeberger’s protocol with slight modifications, comprising 1.5M guanidine thiocyanate and 100mM Tris with 0.1% sodium dodecyl sulfate (SDS) at pH 8.25 Concentration of DNA ranged from 0.05 ng/uL to 6.03 ng/uL whereas the ratio obtained from analysis of DNA purity (260 nm/280 nm) ranged from 1.4 to 2.17. The DNA extracts were stored at -20°C and used for PCR amplification.
Outer and nested PCR amplification was conducted using the AB 9800 Fast Thermocycler machine (Applied Biosystems, UK) on regions flanking identified point mutations of the following P. falciparum genes: bifunctional dihydrofolate reductase-thymidylate synthase – DHFR-TS (PF3D7_0417200), i.e. N51I, C59R, and S108N, hydroxymethyldihydropterin pyrophosphokinase-dihydropteroate synthase – PPPK-DHPS (PF3D7_0810800) i.e. K540E and kelch protein – kelch 13 (PF3D7_1343700) i.e. Y493H, R539T, I543T, and C580Y which confer drug resistance to SP and ART respectively. The respective gene sequences were retrieved from PlasmoDB release 46 (http://PlasmoDB.org) and primer design (Table 1) was performed using PrimerQuest and OligoAnalyzer tools from Integrated DNA technologies online platform (https://www.idtdna.com/). Selection of primers considered characteristics such as: Guanine-Cytosine (G+C) content of greater than 50, five degrees difference between melting temperatures and absence of hair-pin formation and self-annealing properties. A total PCR volume of 25 uL containing 12.5 uL of 2× DreamTaq PCR master mix (Thermo ScientificTM), 3.75 uL of the DNA template and 0.25 uL of the forward and reverse primers respectively were obtained for all the reactions. A volume of 3.75 uL of DNA template in the outer primary PCR reaction, as well as for the PCR amplicon in the nested secondary reaction was used. Step-down PCR cycling conditions for the outer and nested reactions were set as follows: an initial denaturation of 94°C for three minutes, a denaturation of 94°C for 15 seconds, an annealing temperature range of 55°C–60°C for 30 seconds, an elongation of 72°C for one minute and a final elongation of 72°C for 10 minutes.
Resolution of PCR amplicons was run in 1.5% agarose gel, 1× TAE buffer, at 70 V, 58 mA for one hour 30 minutes using a gel electrophoresis system (IBI-Shelton Scientific MP-1015 multipurpose) and an electrophoresis power voltage supplier (Pharmacia LKB ECPS 3000V/150mA). GelRed® Nucleic Acid Gel stain (Biotium) was used for pre-cast gel staining, 1 kb DNA ladder (Thermo ScientificTM) for DNA quantification of resolved PCR amplicons. P. falciparum 3D7 purified DNA laboratory strain obtained from Kenya Medical Research Institute (KEMRI) was used as the main control for wild-type and mutant alleles of each gene. Purification of nested PCR amplicons depicting a single band was performed using the QIAquick PCR purification kit (Qiagen) whereas for amplicons showing double bands, the targets were processed using QIAquick gel extraction kit (Qiagen) as per the manufacturer’s protocol respectively. The PCR amplicons were shipped to Macrogen (Seoul, Korea) for Sanger sequencing.
QIAGEN CLC Main Workbench v21.0.4 was used to perform sequence data editing, consensus sequence assembly and identification of nucleotide base conflicts against the 3D7 reference gene sequences of PF3D7_0417200, PF3D7_0810800 and PF3D7_1343700. Multiple sequence alignment (MSA), was carried out in MEGA v7.026 using the Muscle algorithm27 to identify nucleotide base changes, including translation to amino acid sequences using the standard genetic code for the identification of amino acid changes and their respective positions. Further visualisation of sequence alignments was performed in Jalview v2.11.1.428 to identify non-synonymous point mutations.
Microscopic examination of samples collected from the 19 patients (AQH=10; KH=3; GBH=6) on D3 of follow-up, detected P. falciparum parasites in: 6 of the 10 patients (60%) from AQH, 1 of the 3 patients (33.3%) from KH, and 1 of the 6 patients (16.6%) from GBH (Table 2). These eight (8) patients who tested positive for microscopy were re-administered with Quinine for three days and on D7 of follow-up, they did not present asexual parasites (Table 2). For the other remaining 11 patients, no asexual parasites were observed microscopically for every 200 leucocytes counted (Table 2). Rapid diagnostic testing of P. falciparum histidine-rich protein 2 produced a similar number of patient results as microscopy.
On PCR amplification of targeted gene regions, sequence data from eight samples (AQH = 2, KH = 2, GBH = 4) was eventually analyzed for point mutations (Table 3). The nucleotide base changes comprised of four Pfdhfr substitutions, adenine (A) to cytosine (C) at position 152, thymine (T) to cytosine (C) at position 175, guanine (G) to adenine (A) at position 323, thymine (T) to cytosine (C) at position 134; two Pfdhps substitutions, adenine (A) to guanine (G) at position 1618 and 1347 and none identified for PfK-13 (Table 4). Subsequent translation to amino acid sequences constituted changes as follows: asparagine (N) to isoleucine (I) at codon 51, cysteine (C) to arginine (R) at codon 59, serine (S) to asparagine (N) at codon 108 and valine (V) to alanine (A) at codon 45 for Pfdhfr; lysine (K) to glutamate (E) at codon 540 and arginine (R) retained at codon 449 for Pfdhps and wild-type amino acids retained for Pfkelch-13 (Table 4). Multiple sequence alignment (MSA) and visualization of consensus sequence assemblies for Pfdhfr, Pfdhps and Pfkelch-13 against their 3D7 reference sequences distinguished four non-synonymous (nsy) point mutations for Pfdhfr (N51I, C59R, S108N, V45A), one non-synonymous (nsy) point mutation (K540E) and one synonymous (sy) point mutation (R449R) for Pfdhps while Pfkelch-13 retained wild-type amino acids (Figure 2).
N = asparagine, I = isoleucine, C = cysteine, R = arginine, S = serine, V = valine, A = alanine.

Jalview visualization of multiple sequence alignments depicting nsy- and sy-point mutations: Pfdhfr (N51I, S108N, V45A) occurred in all four isolates (KH013, GBH017, AQH010, AQH009), C59R was identified in one isolate (KH013); Pfdhps (K540E) occurred in all four isolates (KH013, GBH017, AQH010, AQH009), R449R was identified in one isolate (GBH015) and Pfdhps (K540E) occurred in all four isolates (KH012, GBH014, GBH015, GBH017) PfK-13, established no point mutations in all six isolates, wild type amino acids retained at c.554(S), c.566(V), c.569(A), c.578(A), c.580(C), c.493(Y), c.539(R), c.543 (I).
Analyses of Pfdhfr, Pfdhps and Pfkelch-13 haplotype combinations identified the following: Pfdhfr triple mutant (S108N, C59R, and N51I), Pfdhfr double mutant (N51I, S108N), Pfdhps single mutant (K540E), Pfdhfr + Pfdhps mixed mutant (S108N, N51I + K540E), and Pfkelch-13 wild types. The number of isolates and distribution of haplotypes according to hospital sites is illustrated on Table 5.
Eritrea is situated in the uppermost limit of malaria distribution in East Africa, resulting in the seasonal and focal nature of transmission in the country.10 In addition to differences in topography and altitude, disease transmission is highly influenced by rainfall and temperatures.29,30 Unlike a majority of SSA where malaria burden is higher in target populations of pregnant women and children under five years, malaria risk in Eritrea is evenly distributed across all age-groups.10 Data from our study showed a higher number of cases in the over 20 years’ group (n=79) than the under 5 years (n=4) across the hospital sites.24 Also our study found more male cases (n=96), than female (n=35), were admitted across the hospital sites.24 This finding is similar to a previous study in Eritrea which reported a higher risk of parasitemia in males than females.10 Despite this, Eritrea is one of four countries globally which has considerably reduced malaria transmission through a combination of vector and parasite management, as well as, community-led awareness campaigns.31–33 P. falciparum, generally has the highest transmission intensity in SSA and Eritrea, where it causes the most severe form of malaria.34,35 Molecular surveillance of point mutations associated with resistance to previous and current antimalarial drugs, is important in strengthening existing control efforts and complementing therapeutic efficacy studies.36,37
In this study, we present findings from a pilot survey assessing the occurrence of point mutations in PfK-13, Pfdhfr and Pfdhps genes from clinical isolates obtained from three zobas of Eritrea: Adi Quala (Adi Quala Hospital), Debub (Keren Hospital) and Anseba (Gash Barka Hospital). We targeted PCR-amplification of PfK-13 point mutations associated with artemisinin (ART) resistant phenotype in western Cambodia, South-East Asia Y493H, R539T, I543T, C580Y,18 nsy point mutations, V566I, A578S, identified in isolates from five Sub-Saharan countries38 and N554S, A569S reported in a previous study from islands in Lake Victoria, Kenya.39 This study aimed at genotyping for ART resistance on D3 after treatment administration since, conventionally, the proportion of parasitemia on D3 is used in monitoring of therapeutic efficacy to determine a likely occurrence of P. falciparum artemisinin resistance.40 We also aimed to identify whether the other nsy point mutations not associated with ART resistance had spread into Eritrea. The A578S and V566I mutations targeted in our study, have been shown to have a prevalence of >1% in various sites of SSA including Kenya, Uganda, Democratic Republic of Congo, Ghana, Mali and Gabon.41 A recent review has reported A578S is most common in SSA, with a prevalence of up to 11% in 14 countries.42 Also prospective to our study, another PfK-13 mutation R622I, first reported in Ethiopia,43 has later been seen to spread into Somalia,44 Eritrea,45 Uganda,46 and Nigeria.47 However, from our study, none of the corresponding point mutations in PfK-13, were detected. This is similar to other studies from Eritrea48 and Kenya49,50 including other malaria endemic sub-Saharan countries.51 Microscopy data on D3 after treatment administration with the prescribed artemisinin (Artesunate [AS]) indicated parasite clearance for 11 patients who were sampled. This corresponded with our genotyping findings, which were negative for resistance-associated point mutations for five of the 11 samples (Table 3), suggesting a likely absence of ART resistance. One isolate (GBH014), obtained from a patient with parasitological evidence on D3 also did not have resistance-associated point mutations in the PfK-13 marker. This treatment outcome could be attributed to other possible causes of treatment failure such as non-compliance to the treatment regimen, incorrect drug usage, drug pharmacokinetics as well as host immunity.52,53
The triple mutant Pfdhfr haplotype (N51I, C59R and S108N) observed in our study correspond with previous reports from Senegal,54 South Africa,55 Malawi, Mali, Kenya, Tanzania,56,57 including Venezuela in South America.58 This triple mutant haplotype has been associated with in vivo SP treatment failure in previous studies.59–61 Additionally, the single mutant Pfdhfr C59R and single mutant Pfdhps K540E point mutations seen in our findings, have been shown to predict the occurrence of the Pfdhfr-Pfdhps quintuple mutant haplotype (Pfdhfr 51I/59R/108N + Pfdhps 437G/540E),62 which is associated with fully resistant SP parasites16 as well as in vivo SP treatment failure.63 The Pfdhps K540E mutation has been reported to occur together with the A437G mutation in East Africa, and they both play an important role in sulfadoxine resistance of African parasites.64 The detection of these Pfdhfr-Pfdhps mutations from our findings, could be attributed to the development of resistance from prior use of the CQ-SP combination as first-line treatment for clinical management of malaria in Eritrea.23 Additionally, prior evidence from genotyping microsatellite loci of Pfdhps and Pfdhfr genes, shows that SP resistant parasites originated from South East Asia and consecutively spread into Sub-Saharan Africa,65,66 which eventually reached Eritrea too, as demonstrated in these findings. The valine (V) to alanine (A) change at codon 45 in Pfdhfr from this study, has not been previously reported, although, a converse occurrence of alanine (A) to valine (V) at codon 16 has been associated, both singly and doubly in combination to S108N mutation, with resistance to another antifolate, cycloguanil.67,68 Further genotyping, in vitro, and genetic transformation studies could be carried out, firstly, to validate the selection of the V45A mutation in the population, and later to understand its implications to protein function in association with other established Pfdhfr mutations. Also, detection of other SP, resistance associated mutations, Pfdhfr I164L, and Pfdhps A581G, A613T/S is recommended to describe the prevalence of parasite resistance in the population.
A limitation of this study was that, PCR amplification did not occur for some samples. This could be attributed to low genomic DNA yield, as well as, storage length and conditions of the DBS samples.69,70 Nonetheless, the general findings reported here, are not affected by these limitations and essentially provides useful molecular information for further studies.
In this study neither the validated point mutations associated with ART resistance nor the other nsy mutations were detected in the PfK-13 genetic marker. However, the single mutant Pfdhfr haplotype C59R, single mutant Pfdhps haplotype K540E, double mutant Pfdhfr haplotype (N51I, S108N), and mixed mutant Pfdhfr-Pfdhps haplotype (S108N, N51I + K540E) were detected and indicate the possible occurrence of the quintuple mutant haplotype (Pfdhfr N51I/C59R/S108N + Pfdhps A437G/K540E) associated with full SP resistance and in vivo SP treatment failure. The Pfdhfr V45A mutation identified here, has not been previously reported, and further studies could be done to validate its selection and assess its contribution to antifolate resistance. Continued monitoring of artemisinin resistance is required to track resistance-associated point mutations arising at the genetic level. Future studies can be carried out on a larger sample size to determine the mutational prevalence of SP resistance.
This project contains the following underlying data:
NCBI Gene: bifunctional dihydrofolate reductase-thymidylate synthase (DHFR-TS) [Plasmodium falciparum (malaria parasite)] Accession number MZ322415, https://www.ncbi.nlm.nih.gov/nuccore/MZ322415.
NCBI Gene: bifunctional dihydrofolate reductase-thymidylate synthase (DHFR-TS) [Plasmodium falciparum (malaria parasite)] Accession number MZ322416, https://www.ncbi.nlm.nih.gov/nuccore/MZ322416.
NCBI Gene: bifunctional dihydrofolate reductase-thymidylate synthase (DHFR-TS) [Plasmodium falciparum (malaria parasite)] Accession number MZ322417, https://www.ncbi.nlm.nih.gov/nuccore/MZ322417.
NCBI Gene: bifunctional dihydrofolate reductase-thymidylate synthase (DHFR-TS) [Plasmodium falciparum (malaria parasite)] Accession number MZ322418, https://www.ncbi.nlm.nih.gov/nuccore/MZ322418.
NCBI Gene: hydroxymethyldihydropterin pyrophosphokinase-dihydropteroate synthase (PPPK-DHPS) [Plasmodium falciparum (malaria parasite)] Accession number MZ322419, https://www.ncbi.nlm.nih.gov/nuccore/MZ322419.
NCBI Gene: hydroxymethyldihydropterin pyrophosphokinase-dihydropteroate synthase (PPPK-DHPS) [Plasmodium falciparum (malaria parasite)] Accession number MZ322420, https://www.ncbi.nlm.nih.gov/nuccore/MZ322420.
NCBI Gene: hydroxymethyldihydropterin pyrophosphokinase-dihydropteroate synthase (PPPK-DHPS) [Plasmodium falciparum (malaria parasite)] Accession number MZ322421, https://www.ncbi.nlm.nih.gov/nuccore/MZ322421.
NCBI Gene: hydroxymethyldihydropterin pyrophosphokinase-dihydropteroate synthase (PPPK-DHPS) [Plasmodium falciparum (malaria parasite)] Accession number MZ322422, https://www.ncbi.nlm.nih.gov/nuccore/MZ322422.
NCBI Gene: kelch protein (K13) (Kelch13) [Plasmodium falciparum (malaria parasite)] Accession number MZ322423, https://www.ncbi.nlm.nih.gov/nuccore/MZ322423.
NCBI Gene: kelch protein (K13) (Kelch13) [Plasmodium falciparum (malaria parasite)] Accession number MZ322424, https://www.ncbi.nlm.nih.gov/nuccore/MZ322424.
NCBI Gene: kelch protein (K13) (Kelch13) [Plasmodium falciparum (malaria parasite)] Accession number MZ322425, https://www.ncbi.nlm.nih.gov/nuccore/MZ322425.
NCBI Gene: kelch protein (K13) (Kelch13) [Plasmodium falciparum (malaria parasite)] Accession number MZ322426, https://www.ncbi.nlm.nih.gov/nuccore/MZ322426.
NCBI Gene: kelch protein (K13) (Kelch13) [Plasmodium falciparum (malaria parasite)] Accession number MZ322427, https://www.ncbi.nlm.nih.gov/nuccore/MZ322427.
NCBI Gene: kelch protein (K13) (Kelch13) [Plasmodium falciparum (malaria parasite)] Accession number MZ322428, https://www.ncbi.nlm.nih.gov/nuccore/MZ322428.
NCBI Gene: kelch protein (K13) (Kelch13) [Plasmodium falciparum (malaria parasite)] Accession number MZ322429, https://www.ncbi.nlm.nih.gov/nuccore/MZ322429.
NCBI Gene: kelch protein (K13) (Kelch13) [Plasmodium falciparum (malaria parasite)] Accession number MZ322430, https://www.ncbi.nlm.nih.gov/nuccore/MZ322430.
NCBI Gene: kelch protein (K13) (Kelch13) [Plasmodium falciparum (malaria parasite)] Accession number MZ322431, https://www.ncbi.nlm.nih.gov/nuccore/MZ322431.
Dryad: Extended data for ‘Screening for Antifolate and Artemisinin resistance in Plasmodium falciparum clinical isolates from three hospitals of Eritrea’, https://doi.org/10.5061/dryad.sbcc2fr6q.24
This project contains the following extended data:
• the total number of patients grouped according to age, who visited the three hospitals during the study period.
• gel images of Pfdhfr, Pfdhps and PfK13 genetic markers.
Data are available under the terms of the Creative Commons Zero “No rights reserved” data waiver (CC0 1.0 Public domain dedication).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)