Keywords
in vitro model, miniaturisation, animal reduction, high throughput, anticoccidial compounds, chicken coccidiosis, Eimeria species
This article is included in the NC3Rs gateway.
in vitro model, miniaturisation, animal reduction, high throughput, anticoccidial compounds, chicken coccidiosis, Eimeria species
The new version of the manuscript has included all the recommendations done by the reviewers. It also provides new material (including two new figures) and new statistical analysis to support authors’ new findings and claims.
See the authors' detailed response to the review by Benjamin Adjei-Mensah
See the authors' detailed response to the review by Joseph D. Turner
Scientific benefit:
• Increased throughput for the evaluation of new compounds aimed to control chicken coccidiosis by miniaturising the previous in vitro compound-screening model
3Rs benefit:
• Evaluation of novel anticoccidial compounds will require fewer chickens for the generation of parasite stocks
Practical benefits:
• Reduced variability: simultaneous evaluation of a larger number of anticoccidial compounds
• Cost effectiveness: less materials and reagents are necessary per evaluated compounds
• Streamlined protocols: 96-well plate format is used through the whole process (cell culture, DNA isolation and qPCR analysis)
Current application:
• High-throughput screening of potential anticoccidial compounds under investigation by commercial companies and/or public research organisations
Potential applications:
The use of in vitro models in biosciences and biomedical research has supported many scientific advances and has emerged as a suitable alternative to reduce time and cost associated to in vivo models by replacing and/or reducing the use of experimental animals in many areas. The implementation of in vitro models for anticoccidial drugs pre-screening as an alternative to in vivo tests has led to the replacement and reduction of an important number of chickens used in research in coccidiosis control (Thabet et al., 2017). Chicken coccidiosis is an enteric disease caused by different species of the genus Eimeria (Apicomplexa) (Burrell et al., 2020), characterized by malabsorption, diarrhoea and haemorrhage with an important impact on chicken meat and egg production worldwide, estimated in >£10 billion per annum losses (Blake et al., 2020). Coccidiosis has also impact on the ‘Five Freedoms’ of animal welfare, more in particular affecting the ‘freedom from discomfort’ and ‘freedom from pain, injury’ (Webster, 2001). Resistance against current anticoccidial drugs, as well as the near-prohibition of ionophores antibiotics in some parts of the world and public concerns about the use of drugs in animals for human consumption, has led to an increased research in new substitute compounds, evidenced by the high number of new publications in the area in recent years (>70 in vivo studies within the past two years involving >32,000 chickens).
We previously published an in vitro model to evaluate anticoccidial compounds effects in Eimeria tenella ( Marugan-Hernandez et al., 2020); its use for the pre-screening of two essential oils (garlic and oregano) (Sidiropoulou et al., 2020) led to the reduction of the number of chickens used for in vivo testing. The results obtained by the use of the in vitro model showed that each of the essential oils exhibited antiparasitic effects at all the different tested concentrations. This supported the selection of a single experimental group (90 animals + control group) to evaluate a combined treatment of both essential oils at a single dose, excluding the evaluation of the compounds separately, since the in vitro model had already proved their dose-independent individual effects. The in vivo experiment was reduced from six experimental groups to only two (test and control), resulting on a reduction of 90×4=360 chickens in a single study (67%). This in vitro model was standardised and applied using a 24-well plate format (Marugan-Hernandez et al., 2020). In this new study, we aimed to transfer the model to a 96-well plate format. This miniaturisation will reduce the number of parasites needed for compound testing, and therefore the number of chickens used to generate parasites stocks. Additionally, this 96-well plate format will allow the evaluation of a higher number of compounds simultaneously. Eimeria spp. cannot complete their life cycle in vitro, therefore, the natural host (chicken) is necessary to produce stocks for in vitro experimental procedures. The use of one chicken can generate enough parasite material (oocysts) for the pre-screening of two compounds in the 24-well plate format. The use of the 96-well plate format will allow the pre-screening of 10 compounds per animal, this has the potential to reduce local animal use for the study of anticoccidial compounds by 80%.
This study was carried out in strict accordance with the Animals (Scientific Procedures) Act 1986 (United Kingdom Parliament Act). All animal studies and protocols were approved by the Royal Veterinary College Animal Welfare and Ethical Review Board (AWERB) and performed under the United Kingdom Government Home Office under specific project licence (PDAAD5C9D).
Eimeria tenella Wisconsin (Wis) strain (McDougald & Jeffers, 1976) was propagated in ten four-week-old White Leghorn chickens reared under specific pathogen-free conditions. Each chicken was inoculated with 4,000 oocysts to obtain sufficient oocysts for all the in vitro experimental procedures. Table 1 summarises the detailed information of the experiment and used animals following ARRIVE guidelines.
| Study design | |
| Sample size | |
| Inclusion/exclusion criteria | |
| Randomisation | |
| Blinding | |
| Outcome measures | |
| Statistical methods | |
| Experimental animals | |
| Experimental procedures | |
| Results |
Pastor-Fernandez et al. (2019) provides detailed protocols for oocysts isolation and excystation as well as for sporozoite purification. Sporulated oocysts were stored in water at 4 °C for up to six months, the moment from which they start to lose viability for in vitro tests. Freshly purified sporozoites were used to infect cell monolayers immediately.
Standard protocols for cell culture maintenance used here were the same described for the standardisation of the in vitro model for compound-screening (Marugan-Hernandez et al., 2020). Briefly, Madin-Darby bovine kidney (MDBK) cells (NBL-1; ECACC-Sigma-Aldrich) were used as host cell cultures and maintained in a 37 °C humidified incubator with 5% CO2 atmosphere. Cells were cultured in Advanced Dulbecco’s modified Eagle’s Medium (AdDMEM; Gibco) supplemented with 2% heat-inactivated foetal bovine serum (FBS; Sigma) and 100 U/mL penicillin/streptomycin (Fisher, Leicestershire, UK). Confluent cell monolayers (100%) were passaged twice a week to a new confluence of 70-80% by washing them in Ca 2+- and Mg 2+-free PBS and released with 3 mL 0.25% Trypsin/EDTA (Gibco), incubated for three minutes, followed by neutralisation with 5 mL volume of complete medium. The cell suspension was centrifuged at 1,500 g for 10 minutes at room temperature after which the pellet was suspended in 5 mL of fresh medium. Cells were counted and seeded on T75 flasks (Thermo Fisher Scientific) at a density of 10×106 cells with AdDMEM at a final volume of 15 ml or seeded on flat bottom 96-well microtiter plates (Thermo Fisher Scientific Nunc) at a cell density of 0.05×106 cells in each well in 100 μl of AdDMEM culture media for subsequent infection with sporozoites (see Optimisation of infection ratios section).
Optimisation of infection ratios
Based on the amount of 0.3×106 per well in 500 μl, previously optimised for the 24-well plates format to confer a confluence of 100% without excess of cells (where the surface of each well is four time larger than in the 96-well plate format; 0.3×106/4 = 0.075×106), we tested 0.1×106 and 0.05×106 cells per well for the 96-well plate format (just above and below to the equivalent figure). Cells were seeded on flat bottom 96-well microtiter plates (Thermo Fisher Scientific Nunc) at a cell density of 0.1×106 or 0.05×106 cells in each well in 100 μl of AdDMEM culture media for subsequent infection after two hours with sporozoites at different sporozoites:cells ratios (1:1, 2:1, 4:1; Table 2). Three wells were used per conditions (A, B, C) and incubated at 41 °C, 5% CO2. This temperature is necessary for parasite development, as chicken Eimeria parasites develop in the intestine of chickens, whose internal temperature is 41 °C, higher than mammalian hosts. MDBK cells can support this increased temperature for up to six/seven days, longer than the required time for in vitro parasite replication. After 4 hours, infected monolayers were detached with 3 mL 0.25% Trypsin/EDTA (Gibco) and cells counted in a Fuchs-Rosenthal counting chamber. Empty cells were recorded as non-infected, cells with the presence of at least one sporozoites were recorded as infected. Two counts were done per well (Table 2).
| Ratio sporozoites:cells (no. sporozoites) | Wells | Non-infected cells | Infected cells | % Infected cells/count | % Infected cells/well | % Infected cells (average) | tStandard deviation |
|---|---|---|---|---|---|---|---|
| 0.05×106 cells/well | |||||||
| 1:1 (0.05×106) | A | 21 | 16 | 43.2 | 36.1 | 39.0 | 7.6 |
| 27 | 11 | 28.9 | |||||
| B | 26 | 18 | 40.9 | 47.6 | |||
| 21 | 25 | 54.3 | |||||
| C | 26 | 13 | 33.3 | 33.3 | |||
| 22 | 11 | 33.3 | |||||
| 2:1 (0.1×106) | A | 12 | 23 | 65.7 | 60.2 | 60.7 | 7.2 |
| 19 | 23 | 54.8 | |||||
| B | 7 | 21 | 75.0 | 68.1 | |||
| 12 | 19 | 61.3 | |||||
| C | 22 | 21 | 48.8 | 53.8 | |||
| 14 | 20 | 58.8 | |||||
| 4:1 (0.2×106) | A | 7 | 14 | 66.7 | 68.1 | 73.2 | 5.2 |
| 7 | 16 | 69.6 | |||||
| B | 10 | 26 | 72.2 | 73.0 | |||
| 16 | 45 | 73.8 | |||||
| C | 11 | 35 | 76.1 | 78.5 | |||
| 9 | 38 | 80.9 | |||||
| 0.1×106 cells/well | |||||||
| 1:1 (0.1×106) | A | 22 | 30 | 57.7 | 59.5 | 57.3 | 2.0 |
| 22 | 35 | 61.4 | |||||
| B | 21 | 29 | 58.0 | 56.6 | |||
| 22 | 27 | 55.1 | |||||
| C | 26 | 34 | 56.7 | 55.7 | |||
| 34 | 41 | 54.7 | |||||
| 2:1 (0.2×106) | A | 17 | 43 | 71.7 | 63.5 | 61.1 | 7.3 |
| 17 | 21 | 55.3 | |||||
| B | 21 | 51 | 70.8 | 66.9 | |||
| 17 | 29 | 63.0 | |||||
| C | 22 | 34 | 60.7 | 52.9 | |||
| 33 | 27 | 45.0 | |||||
| 4:1 (0.4×106) | A | 15 | 33 | 68.8 | 75.8 | 80.0a) | 4.3 |
| 10 | 48 | 82.8 | |||||
| B | 6 | 28 | 82.4 | 79.9 | |||
| 7 | 24 | 77.4 | |||||
| C | 8 | 28 | 77.8 | 84.3 | |||
| 3 | 30 | 90.9 | |||||
Analytical standard preparations of the classical anticoccidial drugs amprolium hydrochloride (AMP), robenidine hydrochloride (ROB) and salinomycin monosodium hydrate (SAL) were purchased from Sigma (Sigma-Aldrich). Drugs were prepared at 10 mg/mL stock in dimethyl sulfoxide (DMSO; Merck) and dilutions for final working concentrations of 50 μg/ml, 20 μg/mL, 5 μg/mL, and 1 μg/mL were prepared freshly from the stock in AdDMEM just before incubation with sporozoites.
A schematic representation of the experimental design is shown in Figure 1. MDBK cells were seeded in four different 96-well plates (one for each time point to be harvested) at a density of 0.05×106 cells per well and left to settle for two to four hours at 37 °C, 5% CO2. Meanwhile, freshly-purified sporozoites (0.2×106 per well) of E. tenella Wis strain were pre-incubated for 1 h at 41 °C, 5% CO2 with each anticoccidial compound at different concentrations. Untreated pre-incubated sporozoites were used as controls for invasion and replication. After pre-incubation, sporozoites were centrifuged at 1,500 g for 10 minutes at room temperature and washed with PBS to rinse them free of drugs, then resuspended in 300 μl of AdDMEM (to plate 100 μl per well in triplicate). Sporozoites were then added to MDBK monolayers seeded in 96-well plates and incubated at 41 °C, 5% CO2. After two hours, all the wells were washed twice in AdDMEM to remove extracellular sporozoites. The first time point was harvested at two hours post infection (hpi) by washing twice with PBS, then adding RTL buffer (Qiagen) and storing the plate at -20 °C until used for genomic DNA isolation. The same process was done at 24, 44 and 52 hpi. Three wells were used per condition (for untreated and treated sporozoites and for each drug concentration at each time point). Two different biological replicated were performed in different weeks.
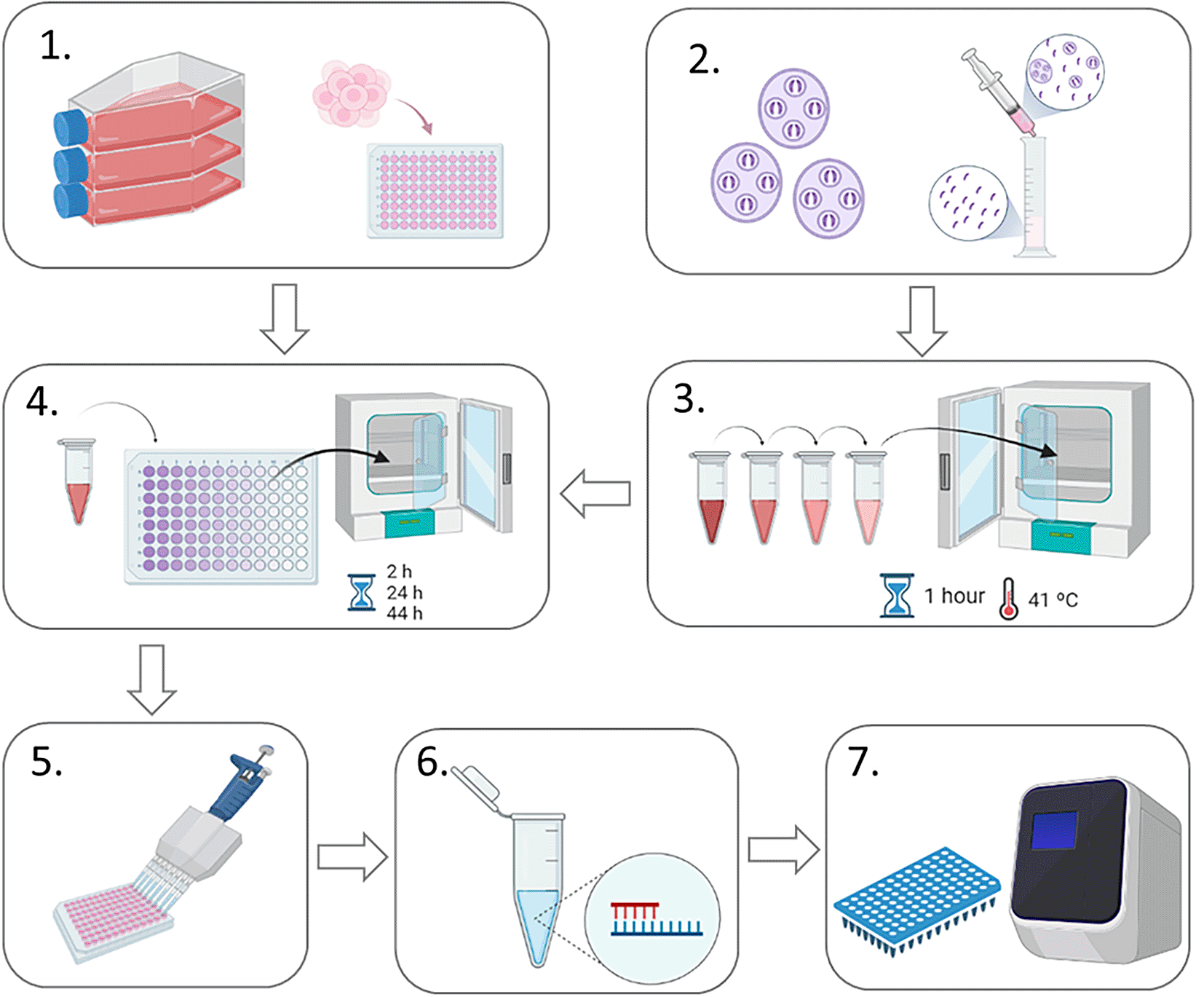
1. MDBK cells were seeded into 96-well plates to form a confluent monolayer (0.05 × 106 cells/well); 2. Sporozoites were purified from oocysts using DE-52 columns (Pastor-Fernandez et al., 2019); 3. Sporozoites were pre-incubated with anticoccidial drugs at four different concentrations (or without drugs for controls) for 1 hour at 4°C; 4. Sporozoites were centrifuged, resuspended in AdDMEM and added to MDBK monolayers to allow invasion. 5. Infected cell monolayers were lysed and collected; 6. Genomic DNA was extracted from collected infected cell monolayers; 7. Real time qPCR was used to evaluate the number of parasite genomes for each sample.
Genomic DNA (gDNA) was isolated from the samples stored in RTL buffer (Qiagen) using the AllPrep DNA/RNA 96 Kit (Qiagen) coupled to the QIAvac 96 (Qiagen) following manufacturer’s instructions.
CFX96 Touch R Real-Time PCR Detection System (Bio-Rad) was used to perform the quantitative PCR following the procedures described previously, using DNA-binding dye SsoFastTM EvaGreen Supermix (Bio-Rad) (Marugan-Hernandez et al., 2016). For parasite quantification, the number of haploid genomes (equivalent to individual sporozoites or merozoites) per well (three wells/sample, technical replicates) was determined for each condition using gDNA specific primers for E. tenella 5S rDNA (Fw_5S: TCATCACCCAAAGGGATT, Rv_5S: TTCATACTGCGTCTAATGCAC) (Clark et al., 2008) and a standard curve of sporozoite gDNA extracted by the same methods (gDNA equivalent to 107 followed by serial dilution to 102). Once the run was completed, a baseline was calculated by Bio-Rad CFX Manager software (Bio-Rad) and applied to each sample to compare the quantification cycles (Cq values) obtained from different wells.
The number of genomes (sporozoites) for each data point analysed by qPCR, was automatically determined by regression analysis using Bio-Rad CFX Manager software (Bio-Rad). Three experimental replicates were included per sample to allow single points showing a standard deviation of more than 0.5 to be excluded from the analysis without affecting the quantification, if all three curves were out of this range, qPCR was repeated for this sample.
Correlation (r) between sporozoite quantification between the 24 and 96-well plate format cultures was determined using the nonparametric Spearman's rank test using GraphPad Prism version 6.00 (GraphPad Software, La Jolla, California, USA).
Comparison of groups evaluating differences in ‘number of zoites’ after anticoccidial drug treatment vs. controls was performed using non-parametric Kruskal-Wallis test. The analysis was followed by Dunnett’s multiple comparisons as a post-hoc test. Statistically significant differences were established using a p<0.05. All analyses were performed using GraphPad Prism version 6.00.
The average inhibition percentage for each anticoccidial drug and concentration was calculated following the equation implemented by Thabet et al. (2017):
Comparison of replication rates after drug treatment were done by calculating slopes of the regression line (m) between 24 and 44 hpi, which were calculated following the universal formula:
For the miniaturisation of the model to a 96-well plate format, we screened out the optimal combination of MDBK cells and E. tenella sporozoites. Best monolayer confluences (100% without excess of cells) were achieved with 0.05×106 cells/well. An optimal rate of invasion and development without causing multiple sporozoite infection per cell was obtained with a 4:1 sporozoites:cell ratio (0.2×106 sporozoites/well). Parasite invasion and development was evaluated in a time course experiment followed by qPCR (Figure 2). Increasing amounts of parasite DNA were detected from 24 hpi onwards as described for the 24-well format, indicative of nuclear replication. The linear rate of replication showed a very strong correlation between the 96 and the 24-well plate format models (r=1; 95% confidence interval; Figure 2), validating in this way the suitability of a 96-well plate format to track and evaluate sporozoites invasion and replication. Higher variability was found for the latest time point (52 hpi) which was explained by the rupture of schizonts which will release merozoites as has been observed before (Marugan-Hernandez et al., 2020); these merozoites are washed from the monolayers and therefore not contributing to the parasite numbers when analysed by qPCR. Fewer parasite number were detected for the 96-well plate format along the time course, which correlated with the lower initial numbers of sporozoites used for monolayers infection (0.2 million/well versus 1 million/well).
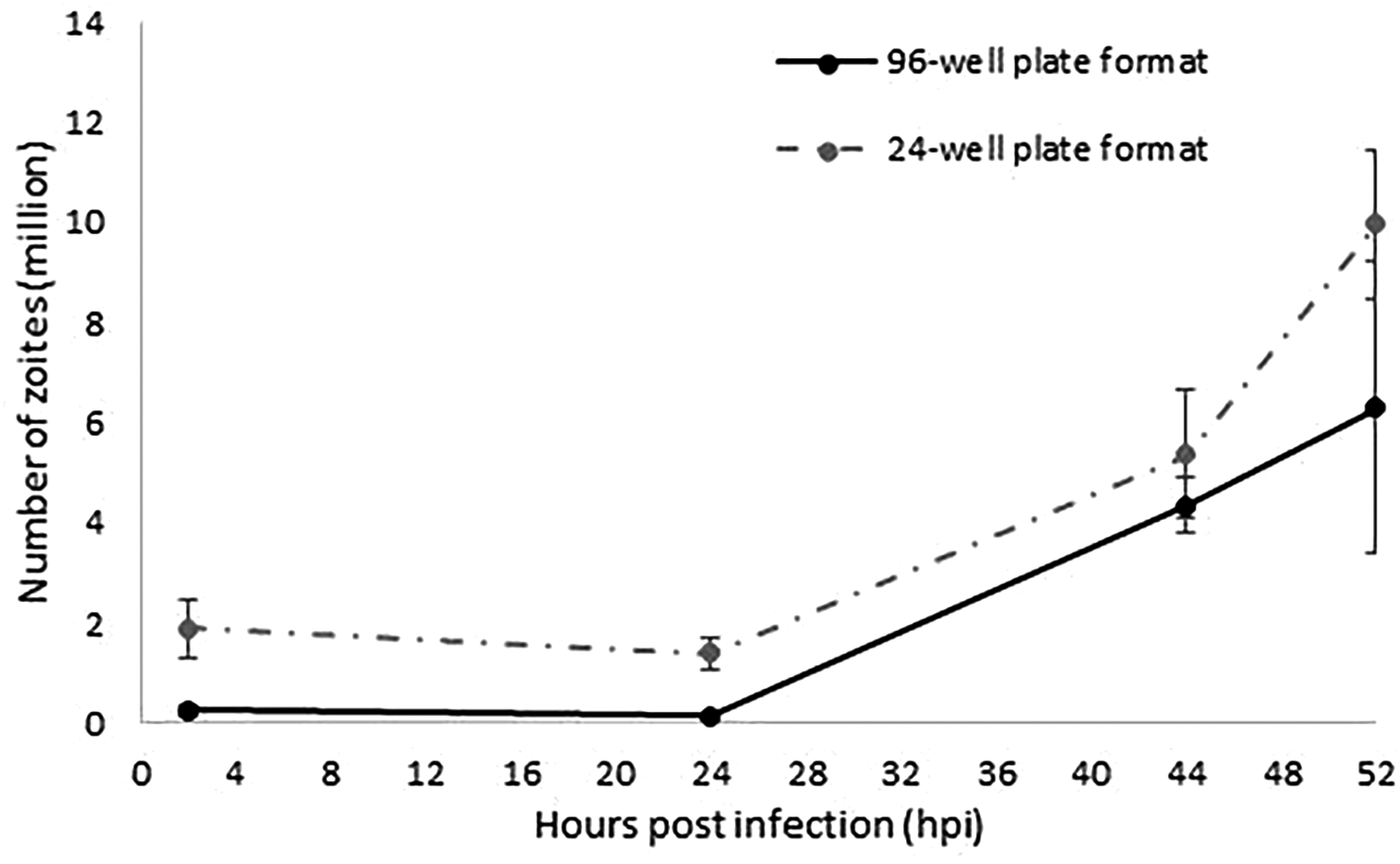
The continuous line represents the number of zoites quantified by qPCR in samples collected at 2, 24, 44 and 52 hpi as the average of the two different replicated experiments performed in separated weeks in a 96-well plate format. Discontinuous line represents data extracted from Marugan-Hernandez et al. (2020) equivalent to the number of zoites quantified by qPCR at the same time points as the average of the three different replicated experiments performed in separated weeks in a 24-well plate format. Error bars represent the standard deviation between repeated experiments for each plate format. Both types of plate format show an equivalent linear fashion growth after 24 hpi (r=1).
Pre-treatment of sporozoites with different concentrations of AMP, ROB and SAL, three drugs with reported anticoccidial activity, showed a relative dose dependant effect (Figure 3). There were no significant differences in the number of sporozoites able to invade host cells after the pre-treated groups vs. the controls at either 2 and 24 hpi (Figure 3A and B). However, treatment with the highest dose of AMP (50 μg/ml) show a significant reduction in replication at both 44 and 52 hpi. Similarly, treatment with ROB (50 μg/ml) exhibited a significant reduction in parasite numbers at 52 hpi (Figure 3C and D).
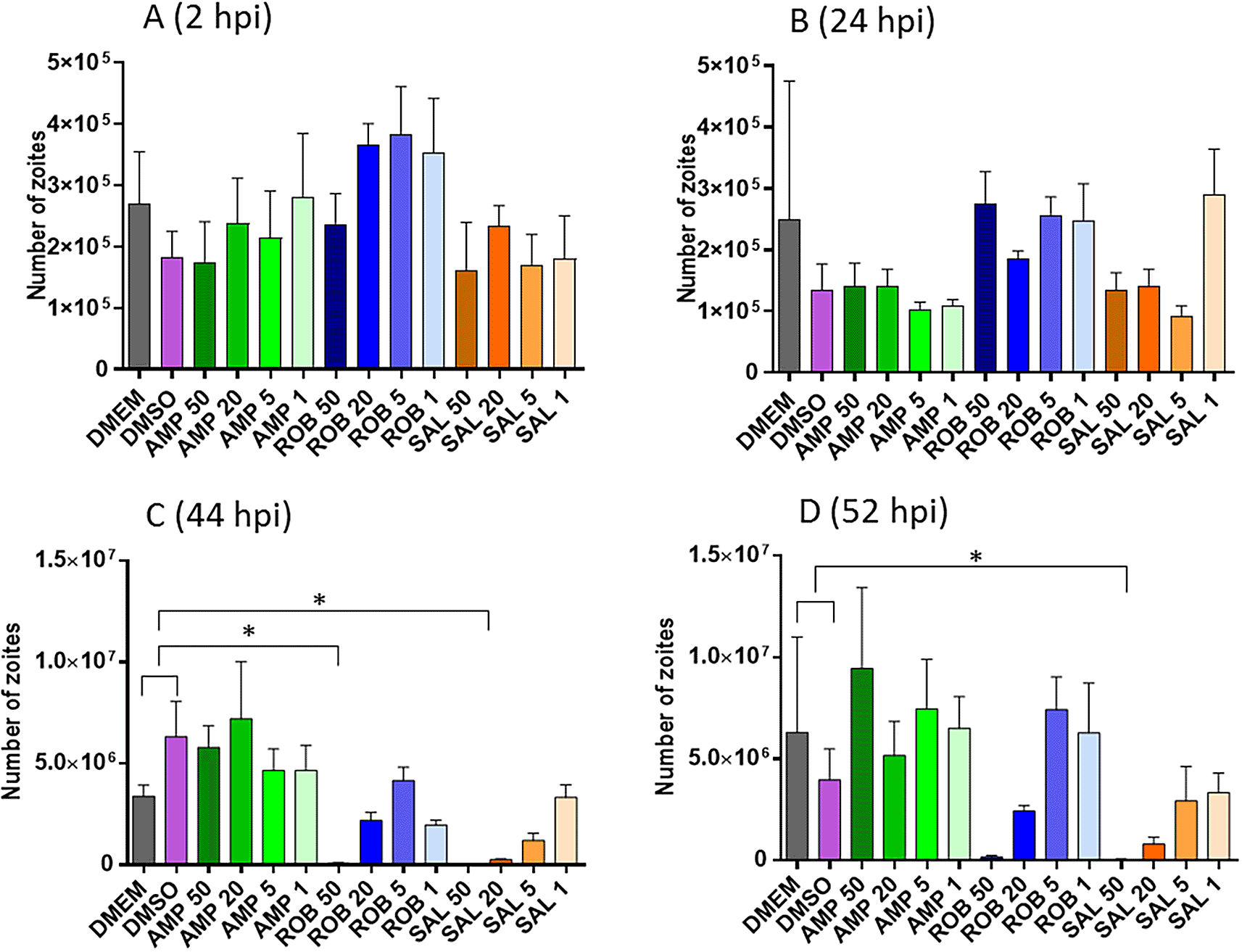
A and B. Graphs shows the number of parasites by qPCR after invasion at 2 and 24 hpi, respectively (before nuclear division has started, see Figure 2); C and D. Graphs shows the number of parasites by qPCR after nuclear replication at 44 and 52 hpi, respectively. X-axis numbers represent the specific drug dilution used in μg/ml. Asterisks indicate significant differences with the DMEM/DMSO controls (Dunnett’s multiple comparison test p<0.05).
Average inhibition of invasion and replication were calculated for each pre-treatment (Table 3). The pre-treatment with AMP at any concentration caused a moderate inhibition of invasion at 2 and 24 hpi (10-45%; average of 22%); however, those sporozoites which were successful at invasion then developed at the same rate as the untreated controls (slope of the regression curve was parallel to that exhibited by the untreated control). Pre-treatment with ROB had little effects on invasion (inhibition of 0-20%, average 5.6%); nevertheless, sporozoites did not replicate well once intracellularly (slope decreased compared with untreated controls, with sporozoite numbers decreasing for the highest concentration, 50 μg/mL). Pre-treatment with SAL exhibited effects in both invasion and development. Invasion was inhibited at higher levels than for AMP (10-60%; average 27%) and intracellular replication was severely impaired for the higher concentrations (50 and 20 μg/mL). In general, for every drug, higher inhibition of invasion and replication was observed for the higher concentration, although some variations were observed.
Each value represents the average of the percentage of inhibition±standard deviation or slope of two different experiments including each three wells per condition, calculated using the equations displayed in Data analysis section.
| Anticoccidial drugs | Concentration (μg/ml) | 2 hpi | 24 hpi | 44 hpi | 52 hpi | Slope (24-44 hpi)* |
|---|---|---|---|---|---|---|
| AMP | 50 | 22.05±14.18 | 7.17±7.17 | 12.88±12.88 | 23.54±23.54 | 282486 |
| 20 | 17.34±10.96 | 10.19±10.19 | 0±0 | 14.11±6.08 | 264669 | |
| 5 | 22.89±4.85 | 24.45±24.45 | 1.96±1.96 | 0±0 | 227343 | |
| 1 | 46.78±5.45 | 25.71±25.71 | 5.13±5.13 | 6.05±6.05 | 227636 | |
| ROB | 50 | 9.13±9.13 | 7.91±7.91 | 97.81±0.07 | 48.49±48.49 | -9013 |
| 20 | 0±0 | 9.07±9.07 | 45.43±23.7 | 40.99±31.24 | 100536 | |
| 5 | 0±0 | 0±0 | 5.42±3.62 | 0±0 | 194893 | |
| 1 | 0±0 | 19.02±19.02 | 52.34±13.91 | 18.23±18.23 | 89050 | |
| SAL | 50 | 62.42±1.79 | 7.94±7.94 | 98.4±0.4 | 99.23±0.09 | -3380 |
| 20 | 16.47±16.47 | 13.5±13.5 | 94.19±0.04 | 87.87±0.94 | 4664 | |
| 5 | 40.02±6.5 | 31.35±14.43 | 73.66±6.83 | 49.41±20.62 | 55507 | |
| 1 | 44.18±21.42 | 0±0 | 26.1±13.58 | 42.16±7.4 | 152050 |
In this study, a model developed to evaluate anticoccidial effects of compounds in vitro, has been successfully miniaturised from a 24-well plate format to a 96-well format. Conditions were refined to create an optimal confluence of the monolayer, supporting invasion rates of >70% in the new format. These conditions have proved to support schizonts and merozoite development as described before in the original model (Marugan-Hernandez et al., 2020). The levels of sporozoites invasion and replication by qPCR in time course experiments in the miniaturised format have shown the same linear growth fashion than described before, and the evaluation of traditional anticoccidial drugs to validate the model has also proved the suitability of this model to detect different levels of inhibition of invasion and/or replication.
When the same anticoccidial drugs (AMP, ROB and SAL) were evaluated in the 24-well plate format, only concentrations of 5 μg/ml were tested (Marugan-Hernandez et al., 2020). Comparing this same concentration in the 96-well plate format, AMP and SAL showed equivalent effects in parasite numbers in invasion and replication for both models. AMP did not show any significant change regarding the controls at any point, whereas SAL did not show differences in invasion, but replication was significantly reduced at 44 hpi (Two-way ANOVA, followed by Dunnett’s post-hoc test). ROB at 5 μg/ml did not correlate with previous results from the 24-well plate model where strong effects were found in both invasion and replication. In the 96-well plate format, ROB did not show any effect in invasion or replication at 5 μg/ml. Interestingly, this concentration has shown inhibitory effects in subsequent studies (Felici et al., 2023) performed in the 96-well plate format, that were equivalent to the 24-well plate format described before. We attributed the lack of significant effects from ROB at 5 μg/ml in the current study to a loss of effect due to continue use of the drug (where stock has gone several freeze-thaw rounds), which we have observed after using it as inhibition control in further studies.
This 96-well plate model has already supported the evaluation of essential oils and their main active compounds (Felici et al., 2023), where the formers have shown similar effects on invasion and replication than ROB and SAL. This is important as research in natural compounds it is essential to replace current anticoccidials for which several resistances have been developed in field strains threatening their efficacy. These latest results evidence the suitability of the miniaturised model to detect inhibition of invasion and/or replication of natural compounds in addition to classical anticoccidal drugs.
Eimeria parasites are self-limited, monoxenous (single-host) parasites, and despite many efforts, there are no efficient in vitro systems supporting continuous replication or the completion of its life cycle; in consequence, research on this species depends on the use of animals. Examination of the literature shows that testing of novel anticoccidial compounds, primarily natural products, is in expansion. Publications in this area have increase from 91 in 2017 to 157 in 2023, and between 2017 and 2023 a total of 57,170 chickens (>8,000 per annum) were used to evaluate anticoccidial effects of novel compounds. In addition to these chickens already mentioned, there are chickens used in unpublished studies from pharmaceutical/nutrition companies or those only providing negative results, numbers of which are harder to discover. Looking specifically at some 2021 publications, if pre-screening with in vitro models had been used by Fu et al. (2021), concentrations could have been adjusted to reduce animals from 630 to 480 (23.8%). Similarly, the use of in vitro models could have excluded one testing group for both Lima et al. (2021) and Herrero-Encinas et al. (2021), reducing animals from 900 to 720 (20%) and 400 to 320 (20%), respectively. If we extrapolate these data (an average of 20% reduction) to the studies published in 2023, the use of our in vitro model could have avoided the experimental use of 4,070 chickens (out of 20,352) globally, or 11,434 (out of 57,170) in the past 7 years.
Nevertheless, chickens are still needed to generate parasite materials to perform in vitro pre-screenings. Therefore, the miniaturisation of our in vitro model will have a direct impact on the reduction of animal use. Testing one compound at four different concentrations (excluding untreated controls) requires 12 million of sporozoites in the 24-well plate format (4 concentrations × 3 wells × 1 million/well) whereas only 2.4 million are necessary in the 96-well plate one (4 concentrations × 3 wells x 0.2 million/well); this means the miniaturised model requires 5 times less parasite material. Efficiency of sporozoite excystation and purification from oocysts is set at 1:1 in our experimental settings. A single chicken can produce an average of 30 million oocysts without causing clinical symptoms. Therefore, with the use of one chicken, we could test at least ten compounds (2.4 million/compound × 10 = 24 million oocysts, plus untreated compounds) with the 96-well format, whereas only two compounds (12 million/compound × 2 = 24 million oocysts, plus controls) could be tested in the 24-well plate format. This supposes a direct 80% reduction in animal use (testing 10 compounds would require 5 chickens in the 24-well plate format versus only 1 in the 96-well plate format, which means 4 less chickens out of 5), for the evaluation of anticoccidial compounds locally. In our laboratory we use an average of 100 chickens per year to maintain parasites stocks with the purposes of evaluating anticoccidial compounds, this number could be reduced to 20 once the miniaturised model is implemented. I addition to this, we also provide parasite materials to collaborators in research institutions and commercial companies at different locations worldwide, this miniaturised model will also have a significant global impact on the number of chickens undergoing a mild severity procedure for the generation of E. tenella oocysts.
Another advantage of this miniaturisation is the high-throughput capacity to test many compounds simultaneously, while reducing materials, reagents and time to perform them. Up to seven compounds could be tested on a single plate per time point (including controls) using <10 mL of culture media, versus only two compounds per plate using volumes of >20 mL per plate in the 24-well plate format. Time is also significantly reduced by a streamlined protocol using 96-well plates and multichannel pipettes throughout the whole experiment, from cell culture to DNA isolation and qPCR analysis. These added benefits will also impact positively in the wider uptake of the model by other researchers since costs and time to achieved results will be significantly reduced.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)