Keywords
Parkinson's disease, PINK1, genetic mutation, mitochondrial phosphorylation, autophagy pathways, oxidative stress
This article is included in the Genomics and Genetics gateway.
This article is included in the Cell & Molecular Biology gateway.
Parkinson's disease, PINK1, genetic mutation, mitochondrial phosphorylation, autophagy pathways, oxidative stress
We are pleased to present this revised review on PINK1-linked Parkinson's disease (PD), significantly improved following constructive peer review.
In response to reviewer feedback, we have transformed the manuscript from a descriptive summary into a critical, forward-looking analysis. Key revisions include: (1) refining the Introduction to explicitly define knowledge gaps, particularly PINK1's emerging role in innate immunity and neuroinflammation via the cGAS-STING pathway; (2) deepening mechanistic discussions to clarify how specific pathogenic variants disrupt Ser65 phosphorylation, blocking recruitment of the primary autophagy receptors OPTN and NDP52; and (3) adding a dedicated section on therapeutic implications, evaluating precision strategies such as kinase amplification with Kinetin and USP30 inhibition.
These revisions position PINK1 not merely as a regulator of mitophagy, but as a critical intersection point between mitochondrial quality control and immune signaling. By integrating contemporary discoveries with translational insights, we offer a framework that we hope will inspire new research directions and accelerate therapeutic development. We are grateful to the reviewers for their rigorous engagement, which has been instrumental in shaping this final version.
See the authors' detailed response to the review by Ryan L Davis
See the authors' detailed response to the review by Eman Ezzeldien Mohamed
See the authors' detailed response to the review by DOMENICO PRATICO
Parkinson’s disease (PD) is the second most prevalent neurodegenerative disorder worldwide, characterized by the progressive loss of dopaminergic neurons in the substantia nigra pars compacta. According to recent epidemiological data, the global prevalence of PD has more than doubled over the past two decades, a trend largely attributed to aging populations.1,2 While the majority of cases are idiopathic, approximately 5–10% of patients exhibit monogenic inheritance. Among these, loss-of-function variants in the PINK1 gene (PTEN-induced kinase 1; HGNC:14581) are established as a primary cause of autosomal recessive early-onset PD (EOPD). Although PINK1-related PD represents a subset of the total patient population, elucidating its role is critical for understanding the mitochondrial quality control failure that characterizes both familial and sporadic forms of the disease.3
The clinical hallmark of PD is the progressive degeneration of dopaminergic neurons within the substantia nigra pars compacta, leading to classic motor symptoms such as bradykinesia, resting tremor, and rigidity.4 However, it is increasingly recognized that a prolonged prodromal phase, characterized by non-motor symptoms including hyposmia, sleep disturbances, and depression, often precedes motor onset by decades.5 In the context of PINK1-linked PD, the clinical phenotype typically manifests as early-onset Parkinsonism, frequently occurring before the age of 45.3 Notably, patients with PINK1 variants often exhibit a slower disease progression and a sustained, robust response to levodopa compared to those with late-onset sporadic PD.6 Understanding these distinct clinical trajectories is crucial for accurate prognosis and personalized patient management.
The PINK1 gene encodes a 581-amino acid protein that is broadly expressed and structurally composed of an N-terminal mitochondrial targeting sequence, a transmembrane domain, and a C-terminal serine/threonine kinase domain.7 While loss-of-function variants in PINK1 are widely recognized for impairing mitochondrial protection against oxidative stress,8 the precise molecular cascade linking these deficits to clinical neurodegeneration remains incompletely mapped. Specifically, there is a critical knowledge gap regarding how PINK1 dysfunction extends beyond canonical mitophagy to affect broader cellular processes such as innate immunity and proteostasis.9 Moreover, despite extensive mechanistic studies, translating these findings into effective therapeutics has proven difficult. Therefore, the objective of this review is to critically examine the multifaceted mechanisms of PINK1-driven pathogenesis—moving beyond classical mitophagy—and to evaluate the translational potential of targeting this pathway for PD intervention.
Under basal physiological conditions, the cellular levels of PINK1 are maintained at an exceptionally low limit through a rapid and constitutive turnover mechanism. The PINK1 precursor is imported into healthy, polarized mitochondria via the translocase of the outer membrane (TOM) and translocase of the inner membrane (TIM) complexes.10 Upon reaching the inner mitochondrial membrane (IMM), the N-terminal mitochondrial targeting sequence (MTS) is first cleaved by the mitochondrial processing peptidase (MPP). Subsequently, the transmembrane domain is cleaved by the rhomboid protease PARL (presenilin-associated rhomboid-like) between Alanine-103 and Phenylalanine-104.11 This proteolytic processing generates a truncated, unstable form of PINK1, which is retro-translocated to the cytosol and rapidly degraded by the ubiquitin-proteasome system via the N-end rule pathway.12 However, this homeostatic cycle is intimately coupled to mitochondrial bioenergetics. When mitochondria sustain damage and lose their membrane potential (Δψm), the TIM23-mediated import of PINK1 is arrested. Consequently, full-length PINK1 stabilizes on the outer mitochondrial membrane (OMM) with its kinase domain facing the cytosol.13 This accumulation promotes PINK1 homodimerization and trans-autophosphorylation at Serine 228 and Serine 402, events that are requisite for maximizing its kinase activity and initiating the downstream recruitment of Parkin.14 Thus, PINK1 functions as a molecular sensor of mitochondrial quality, converting bioenergetic stress into a distinct phosphorylation signal.
The integrity of the mitochondrial quality control system is fundamentally dependent on the precise catalytic activity of PINK1. Consequently, pathogenic variants in the PINK1 gene predominantly result in a loss of function, disrupting the neuroprotective response described in the previous section. Since the initial identification of the G309D missense variant and the W437X truncation in the PARK6 pedigree by Valente et al.,15 over 70 distinct pathogenic variants have been cataloged. Structurally, these variants are not randomly distributed but are heavily clustered within the highly conserved serine/threonine kinase domain (residues 156–509), underscoring the critical importance of kinase activity for neuroprotection.16
Mechanistically, these variants impair PINK1 function through distinct molecular deficits. A significant proportion of missense variants, such as G309D, L347P, and G409V, induce kinase inactivation by destabilizing the ATP-binding pocket or the catalytic loop. This structural compromise abolishes the essential autophosphorylation events (e.g., at Ser228 and Ser402) required for the recruitment of Parkin.17 Beyond catalytic inactivation, protein instability represents another major pathogenic mechanism. Truncation variants, including W437X and Q456X, often result in the rapid degradation of the transcript via the nonsense-mediated decay pathway or produce unstable protein fragments that fail to accumulate on the outer mitochondrial membrane, even under conditions of cellular stress.18 Furthermore, rare variants located in the N-terminal mitochondrial targeting sequence have been observed to interfere with the efficient import of PINK1 into mitochondria, thereby preventing its correct subcellular localization.19
Collectively, regardless of whether the defect is kinetic (kinase dead) or structural (instability), these perturbations converge on a common pathological outcome: the failure to sense mitochondrial depolarization. This sensory defect prevents the initiation of mitophagy, leading to the progressive accumulation of dysfunctional organelles within dopaminergic neurons.
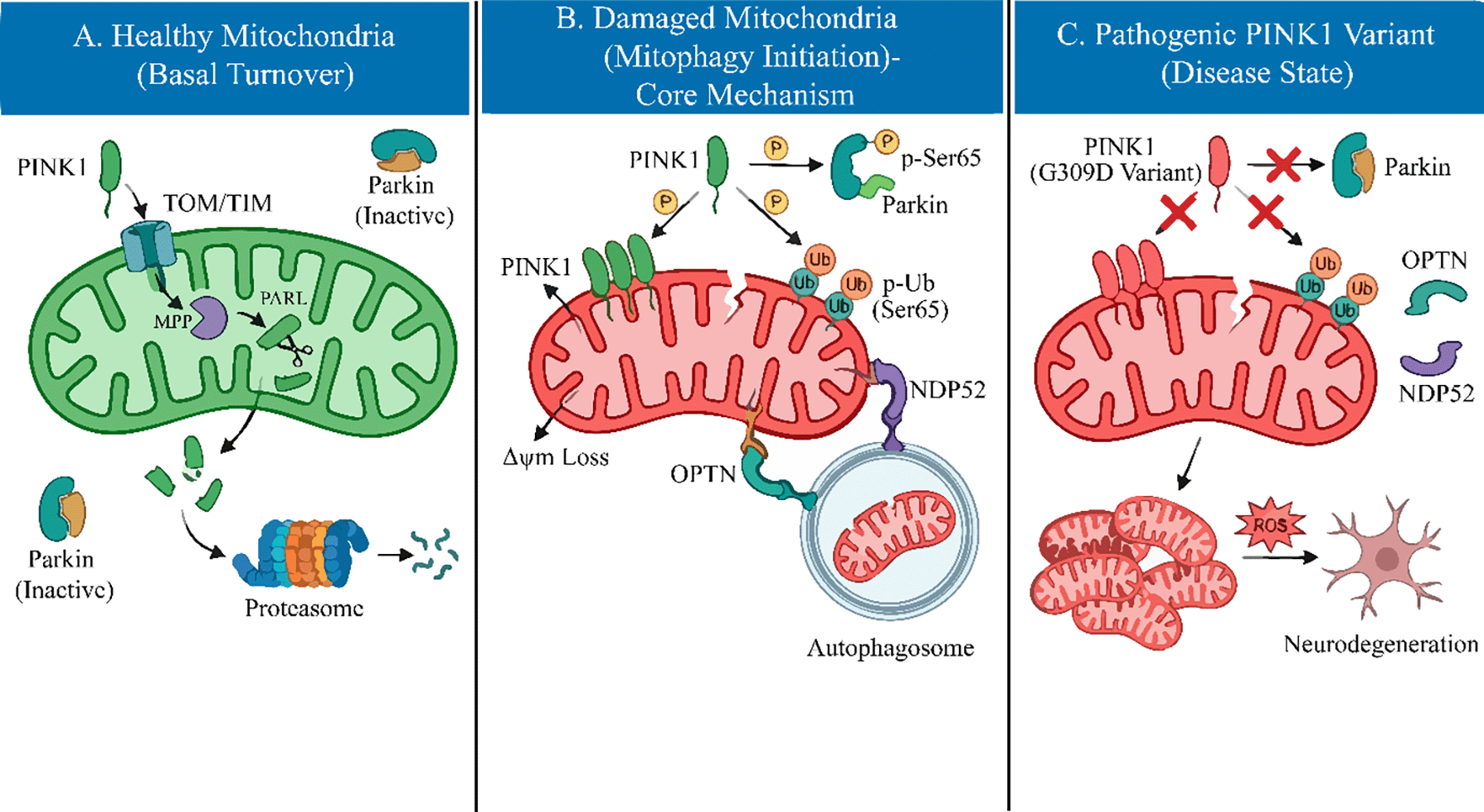
Investigating the interplay between PINK1 kinase activity and its substrates provides crucial insights into PD pathogenesis. Contrary to earlier assumptions that PINK1 broadly phosphorylates mitochondria, it targets specific proteins to mediate neuroprotection. Early studies identified the mitochondrial chaperone TRAP1 and the serine protease HtrA2/OMI as potential substrates. Pridgeon et al. demonstrated that PINK1 phosphorylates TRAP1 to suppress oxidative stress, a function abolished by the G309D variant.8 Similarly, PINK1-dependent phosphorylation of HtrA2 enhances its protease activity, conferring resistance to cellular stress.20
However, the most significant breakthrough has been the identification of Parkin and Ubiquitin as the physiological substrates of PINK1. Under stress, PINK1 phosphorylates both the ubiquitin-like (UBL) domain of Parkin and Ubiquitin itself at the conserved Serine 65 (Ser65) residue.21,22 This dual phosphorylation event is the molecular switch that activates Parkin’s E3 ligase activity (Figure 1). Crucially, pathogenic variants such as G309D and L347P fail to phosphorylate Ubiquitin or Parkin at Ser65, thereby locking Parkin in an auto-inhibited state and completely blocking the downstream quality control cascade.23
Mitophagy, the selective degradation of damaged mitochondria, is the central pathway regulated by PINK1. In the absence of functional PINK1, this clearance mechanism fails. While early models suggested that p62/SQSTM1 was the primary autophagy receptor linking ubiquitinated mitochondria to the autophagosome, recent evidence has redefined this model.
Current consensus indicates that Optineurin (OPTN) and NDP52 (CALCOCO2) are the primary autophagy receptors recruited by PINK1/Parkin-mediated ubiquitin chains.24 These receptors bind to the ubiquitinated outer mitochondrial membrane and recruit the autophagy machinery via their LC3-interacting regions. Importantly, PINK1 further enhances this process by phosphorylating ubiquitin chains, which serves as a “eat-me” signal that selectively recruits OPTN and NDP52.25 Pathogenic PINK1 variants disrupt this recruitment hierarchy. Without the initial phosphorylation trigger from PINK1, Parkin is not activated, ubiquitin chains are not formed, and OPTN/NDP52 cannot engage the autophagy machinery. Consequently, damaged mitochondria accumulate, releasing toxic byproducts that drive dopaminergic neurodegeneration.
Emerging evidence suggests that the consequences of PINK1 dysfunction extend beyond defective mitophagy to include aberrant innate immune signaling. Under physiological conditions, mitophagy prevents the leakage of mitochondrial DNA (mtDNA) into the cytosol. However, in the absence of PINK1, damaged mitochondria accumulate and release mtDNA, which is sensed by the cGAS-STING pathway. This triggers a robust Type I interferon response and the release of pro-inflammatory cytokines such as Interleukin-6 (IL-6).26 Crucially, Matheoud et al. demonstrated that intestinal infection with Gram-negative bacteria in Pink1-knockout mice engages mitochondrial antigen presentation and autoimmune mechanisms, highlighting the gut-brain axis and inflammation as key drivers of pathogenesis.27 Clinically, elevated serum IL-6 levels have been reported in patients with PINK1 or PRKN variants, correlating with disease progression.28 This suggests that PINK1 functions not only as a quality control sensor but also as a critical immunological checkpoint.
Given that PINK1 loss-of-function drives PD, pharmacological strategies to amplify PINK1 activity or bypass its function are being actively explored. One approach involves the neo-substrate Kinetin (N6-furfuryladenine) and its riboside prodrugs, which have been shown to accelerate PINK1 activation and enhance Parkin recruitment in neuronal cells independent of mitochondrial depolarization.29,30 Additionally, inhibitors of the deubiquitinase USP30, which opposes PINK1/Parkin signaling by removing ubiquitin chains from mitochondria, have shown promise in preclinical models by restoring mitophagy flux.31 Recent studies highlight that USP30 inhibition can rescue mitophagy defects even in the presence of certain pathogenic variants, making it a viable therapeutic target.32 These targeted approaches represent the frontier of precision medicine for PINK1-linked Parkinson’s disease.
In summary, this review highlights the intricate relationship between PINK1 loss-of-function variants and the pathogenesis of Parkinson’s disease. The recent identification of Optineurin (OPTN) and NDP52 as the primary autophagy receptors recruited by PINK1-phosphorylated ubiquitin chains has redefined our understanding of mitophagy. Pathogenic variants, by failing to execute the critical Ser65 phosphorylation step, block this recruitment hierarchy, leading to the accumulation of damaged mitochondria. Beyond bioenergetic failure, recent evidence underscores that this defect triggers the cGAS-STING pathway, driving neuroinflammation as a key component of disease progression. A deeper understanding of these mechanisms identifies specific therapeutic targets—such as amplifying PINK1 kinase activity with Kinetin or inhibiting the deubiquitinase USP30—offering promising new avenues for clinical intervention in Parkinson’s disease.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)