Keywords
differentiation, epigenetics, eye development, single-cell ATAC-seq, single-cell RNA-seq
This article is included in the Cell & Molecular Biology gateway.
This article is included in the Japan Institutional Gateway gateway.
differentiation, epigenetics, eye development, single-cell ATAC-seq, single-cell RNA-seq
Signal transduction depends on the expression of receptors regulated at multiple levels. These regulations include chromatin remodeling and DNA-binding proteins, such as transcription factors and transcriptional repressors. Because fine-tuned signal transduction is necessary for development, it is important to clarify the regulatory mechanisms of receptor expression.
Notch signaling in mammals is dependent on the binding of five canonical DSL ligands and four Notch receptors.1 It has been suggested that Notch loci are subject to chromatin remodeling under both normal2–7 and pathological8–16 conditions. In terms of developmental biology, the retina is a good model for investigating cellular differentiation involving Notch signaling.17 The developed retina is composed of multiple cell types, including retinal ganglion cells, bipolar cells, photoreceptors, amacrine cells, horizontal cells, and Müllar glial cells, all of which originate from the retinal progenitor cells (RPCs). RPC is characterized by the expression of Lhx2, Pax6, Rax, and Vsx2 while there are some additional marker genes for developing horizontal cells/retinal ganglion cells (Onecut1/2),18 developing amacrine cells (Elavl2/4),19 developing photoreceptors/amacrine cells/Müllar glial cells (Eef1a1),20 developing retinal ganglion cells (Meis2),21 developing horizontal cells (Lhx1, Ptf1a),22 and glial cells (Pax2).23 Of the four Notch receptors in mammals, Notch1, has been suggested to be involved in the maintenance of RPCs and differentiation into retinal ganglion cells.24 However, it is unclear when and how Notch receptor expression is switched on and off in RPCs during early differentiation. Here, we re-analyzed a public multi-omics dataset of single-cell RNA-seq and single-cell ATAC-seq from three human fetal retinas25 to clarify the timing of Notch receptor expression and examine the involvement of chromatin remodeling in this receptor expression switch.
A single-cell multi-omics dataset (GSE183684)25 was downloaded from the Gene Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo/); data from days 59, 74, and 78 in the dataset were selectively used because they contained many undifferentiated retinal progenitor cells. The data were processed in Seurat version 5.1.026 and Signac version 1.14.027 pipelines in R version 4.4.1 on the Ubuntu 22.04.4 LTS environment. Pseudotime analysis and integration of the single-cell RNA-seq data and the single-cell ATAC-seq data was conducted by using the “FindTransferAnchors” function of Signac and Monocle3 version 1.3.7,28 respectively. The conditions used in the analysis are provided in the GitHub repository.
First, early gestational stage samples were characterized (day 59). Uniform manifold approximation and projection (UMAP) analysis of single-cell RNA-seq identified 13 clusters ( Figure 1A), which were further characterized by marker gene expression ( Figure 1B). This sample primarily contained RPCs with various differentiation statuses except for PAX2-expressing glial cells. Early RPCs (RPC1-3 and MKI67-expressing Proliferating RPC) were characterized by LHX2, PAX6, RAX, and VSX2. In addition, pseudotime analysis suggested that these RPCs differentiated into either ONECUT1/2-expressing, ELAVL2/4-expressing, or EEF1A1-expressing RPCs ( Figure 1C). Next, we examined Notch mRNA expression and found that NOTCH1-3 were expressed primarily in early RPCs, with NOTCH1 and NOTCH3 being the most prominent genes ( Figure 1D). Then, we re-analyzed single-cell ATAC-seq data for this day 59 sample, which was mathematically integrated with its single-cell RNA-seq data by using the “FindTransferAnchors” function of Signac ( Figure 1E). In the early RPCs (RPC1/2 and MKI67-expressing Proliferating RPC), we observed peaks between Chr9 136510000-136520000 of the NOTCH1 gene, which diminished in the other RPC clusters. These results imply that Notch expression transition during RPC differentiation might be associated with the chromatin remodeling of NOTCH1 ( Figure 1F). However, we noted that chromatin accessibility of the upstream region of the NOTCH1 locus remained high in ONECUT1/2-expressing RPCs. In contrast to these populations, chromatin accessibility of the NOTCH1 locus in ELAVL2/4-expressing RPCs was very low, which was consistent with the low mRNA expression in these RPC populations. In addition, the chromatin accessibility of the NOTCH3 locus remained unchanged during RPC differentiation, although its mRNA expression diminished ( Figure 1G). Since NOTCH2 and NOTCH4 were less prominent, the feature plots of the marker genes and the dot plots of the Notch genes from single-cell RNA-seq along with coverage plots of NOTCH2 and NOTCH4 from single-cell ATAC-seq are available in “Additional_file_1” in our GitHub repository.
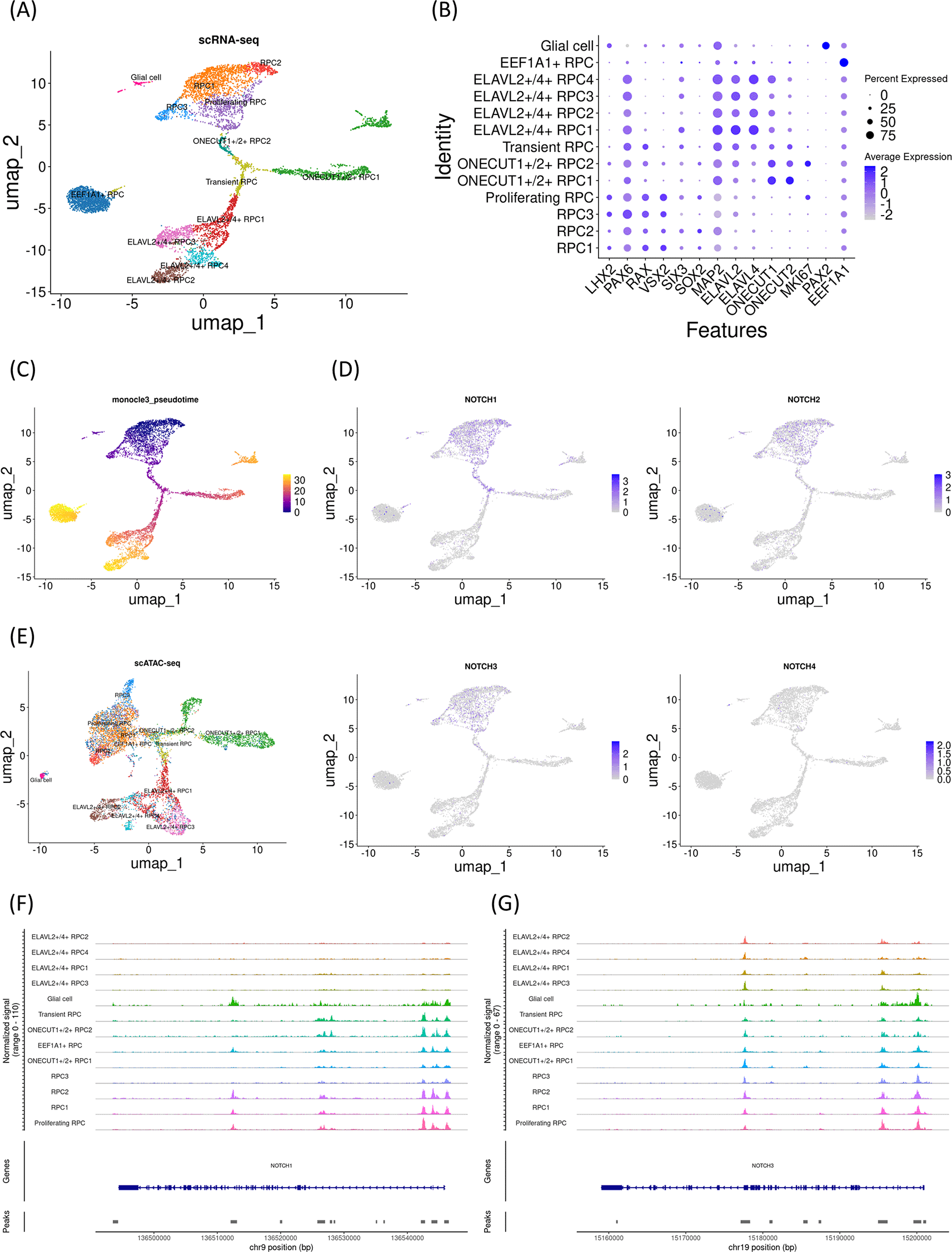
(A) UMAP analysis of the single-cell RNA-seq data identified 13 clusters. (B) Dot plot of marker genes. Note that early RPC clusters expressed LHX2, PAX6, RAX, and VSX2. (C) Monocle3 pseudotime analysis for clarifying the differentiation status. (D) Feature plot of NOTCH1-4. Note that NOTCH1 and NOTCH3 expression was prominent in the early RPC clusters. (E) UMAP analysis of the single-cell ATAC-seq data, which was integrated with the single-cell RNA-seq data. (F) The coverage plot of the NOTCH1 locus showing the chromatin accessibility. Note that the upstream of the gene is on the right. (G) The coverage plot of the NOTCH3 locus. Note that the upstream of the gene is on the right.
To examine whether the changes in Notch mRNA expression and chromatin accessibility of the Notch loci in the day 59 sample were representative, we investigated another early gestational stage sample (day 74). UMAP analysis identified 15 clusters ( Figure 2A) that were further characterized by marker gene expression ( Figure 2B). This sample primarily contained RPCs with various differentiation statuses. Early RPCs (RPC1-6) were characterized by LHX2, PAX6, RAX, and VSX2. In addition, pseudotime analysis suggested that these RPCs differentiated into either ELAVL2/4-expressing, ONECUT1/MEIS2-expressing, or ONECUT2-expressing RPCs ( Figure 2C). ONECUT1/MEIS2-expressing RPCs differentiated into RPCs that markedly expressed VSX2. Next, we examined Notch mRNA expression and found that NOTCH1-3 were expressed primarily in early RPCs, with NOTCH1 and NOTCH3 being the most prominent genes, similar to the day 59 sample ( Figure 2D). We then re-analyzed the single-cell ATAC-seq data for this sample, which were integrated with the single-cell RNA-seq data ( Figure 2E). In the early RPCs (RPC1-3), we observed peaks between Chr9 136510000-136520000 of the NOTCH1 gene, which diminished in the other RPC clusters. These results confirmed that Notch expression transition during RPC differentiation might be associated with the chromatin remodeling of NOTCH1 ( Figure 2F). However, we noted that the chromatin accessibility of the upstream region of the NOTCH1 locus remained high in ONECUT-expressing RPCs. In contrast to these populations, the chromatin accessibility of the NOTCH1 locus in ELAVL2/4-expressing RPCs was very low, which was consistent with the low mRNA expression in these RPC populations. In addition, the chromatin accessibility of the NOTCH3 locus remained unchanged during RPC differentiation ( Figure 2G). In summary, similar to the day 59 sample, in the day 74 sample, a concomitant mRNA decrease and chromatin remodeling in NOTCH1 were observed, while chromatin accessibility in the upstream region of the NOTCH1 locus remained high in ONECUT-expressing RPCs. The feature plots of the marker genes and dot plots of the Notch genes from single-cell RNA-seq along with coverage plots of NOTCH2 and NOTCH4 from single-cell ATAC-seq are available in “Additional_file_2” in our GitHub repository.
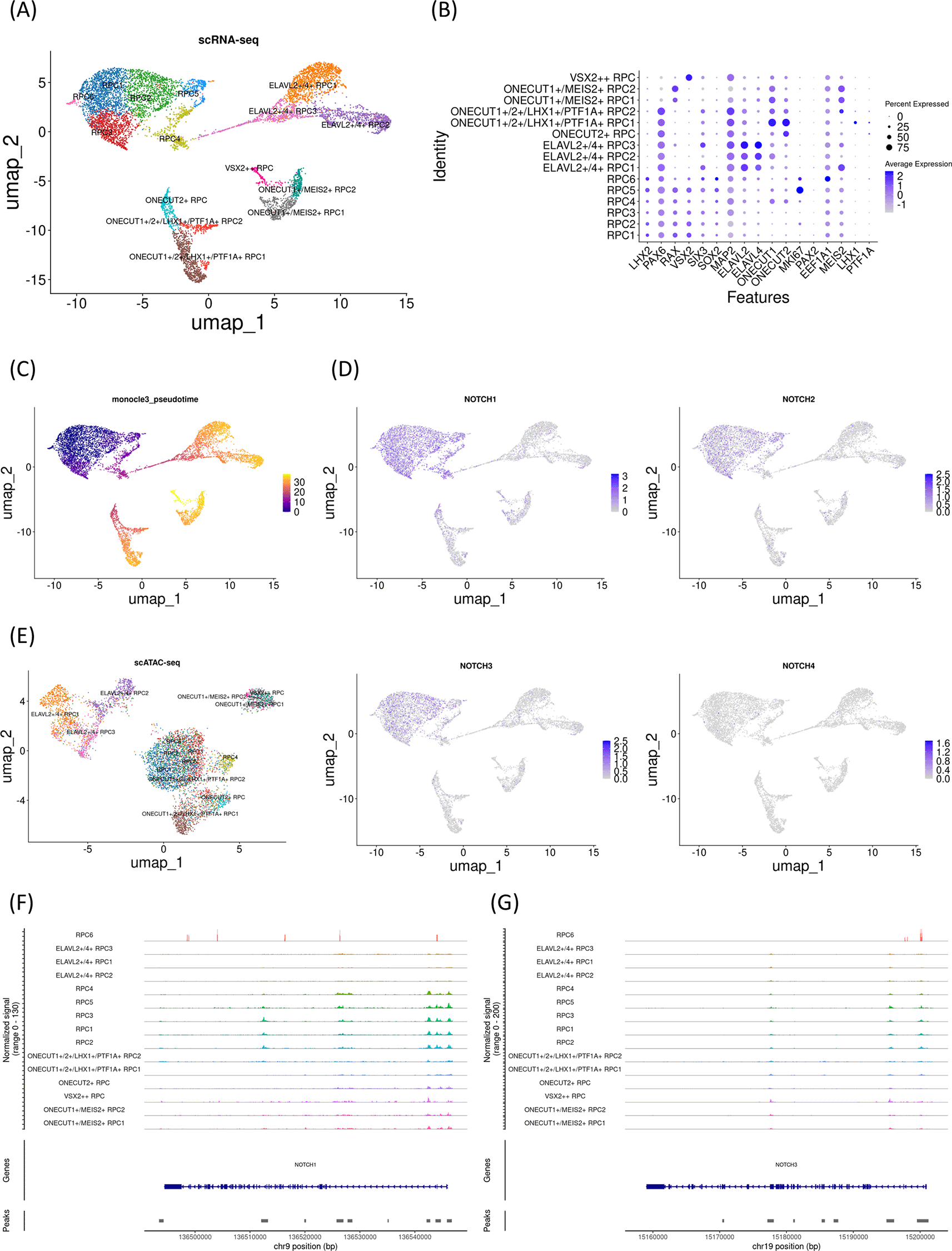
(A) UMAP analysis of the single-cell RNA-seq data identified 15 clusters. (B) Dot plot of marker genes. Note that early RPC clusters expressed LHX2, PAX6, RAX, and VSX2. (C) Monocle3 pseudotime analysis. (D) Feature plot of NOTCH1-4. Note that NOTCH1 and NOTCH3 expression were prominent in the early RPC clusters. (E) UMAP analysis of the single-cell ATAC-seq data which was integrated with the single-cell RNA-seq data. (F) The coverage plot of the NOTCH1 locus. (G) The coverage plot of the NOTCH3 locus.
To further confirm the developmental changes in Notch mRNA expression and chromatin accessibility at the Notch loci, we investigated an additional early gestational stage sample (day 78). UMAP analysis identified 17 clusters ( Figure 3A) that were further characterized by marker gene expression ( Figure 3B). The sample primarily contained RPCs with various differentiation statuses. Early RPCs (RPC1-3 and MKI67-expressing Proliferating RPC1-2 cells) were characterized by LHX2, PAX6, RAX, and VSX2. In addition, pseudotime analysis suggested that these RPCs differentiated into ONECUT1/2-expressing RPCs, which later differentiated into PTF1A and LHX1-expressing RPCs, MEIS2-expressing RPCs, or ELAVL2/4-expressing RPCs ( Figure 3C). Next, we examined Notch mRNA expression and found that NOTCH1-3 were expressed primarily in early RPCs, with NOTCH1 and NOTCH3 being the most prominent genes ( Figure 3D). We then re-analyzed the single-cell ATAC-seq data for the day 78 sample, which was integrated with the single-cell RNA-seq data ( Figure 3E). In the early RPCs (RPC1-3), we observed peaks between Chr9 136510000-136520000 of the NOTCH1 gene, which diminished in the other RPC clusters ( Figure 3F). However, we noted that the chromatin accessibility of the upstream region of the NOTCH1 locus remained high in ONECUT-expressing RPCs. In contrast to these populations, the chromatin accessibility of the NOTCH1 locus in ELAVL2/4-expressing RPCs was very low, which was consistent with the low mRNA expression in these RPC populations. In addition, the chromatin accessibility of the NOTCH3 locus remained unchanged during RPC differentiation ( Figure 3G). In summary, examination of all three independent samples suggested that NOTCH1 mRNA expression decreased, which was concomitant with chromatin remodeling in Chr9 136510000-136520000 of the NOTCH1 locus. The feature plots of the marker genes and the dot plots of the NOTCH genes from single-cell RNA-seq along with coverage plots of NOTCH2 and NOTCH4 from single-cell ATAC-seq are available in “Additional_file_3” in our GitHub repository.
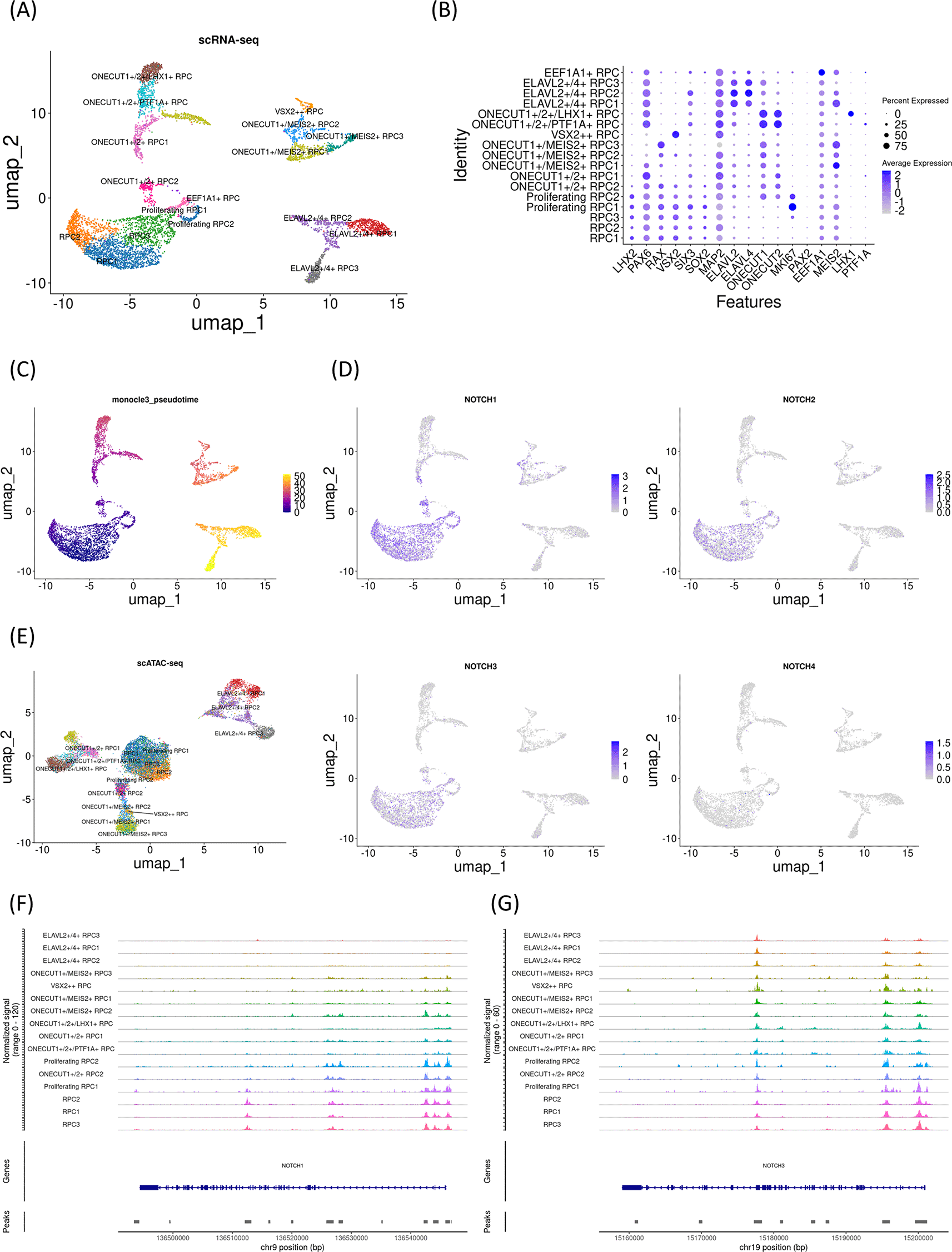
(A) UMAP analysis of the single-cell RNA-seq data identified 17 clusters. (B) Dot plot of marker genes. Note that early RPC clusters expressed LHX2, PAX6, RAX, and VSX2. (C) Monocle3 pseudotime analysis. (D) Feature plot of NOTCH1-4. Note that NOTCH1 and NOTCH3 expression were prominent in early RPC clusters. (E) UMAP analysis of the single-cell ATAC-seq data which was integrated with the single-cell RNA-seq data. (F) The coverage plot of the NOTCH1 locus. (G) The coverage plot of the NOTCH3 locus.
The involvement of Notch signaling in cell fate choices is well documented, including in Drosophila neurogenesis29 and mammalian biliary development.30 Although the regulation of Notch receptor expression is necessary for these processes, to the best of our knowledge, few studies have used genome-wide investigations of the underlying molecular mechanisms. To examine chromatin remodeling in such regulatory mechanisms, we re-analyzed a single-cell RNA-seq and ATAC-seq dataset from developing retinas in which differentiation trajectories were well characterized. By re-analyzing three independent samples, we observed chromatin remodeling in part of the NOTCH1 locus, concomitant with changes in its mRNA expression during RPC differentiation.
Transcriptional regulation occurs at many levels, including DNA-binding proteins and miRNAs, as well as chromatin remodeling. Importantly, the chromatin accessibility of the upstream regions of the NOTCH1 locus was unaffected in ONECUT-expressing RPCs. Therefore, the observed mRNA changes might be driven by pathways other than chromatin remodeling. Indeed, NOTCH3 mRNA expression also diminished during differentiation, although we observed no chromatin remodeling in the NOTCH3 locus. Because chromatin accessibility in ELAVL2/4-expressing RPCs was very low, this ensured low mRNA expression of NOTCH1.
An ophthalmological study revealed that the epigenetic landscape of cell type-specific enhancers shifted during differentiation of RPCs.31 For example, in the single-cell ATAC-seq data from embryonic day 14.5 mouse retina, motif enrichment for Lhx2, Rax and Pax6 in the early RPCs were observed, and footprinting analysis validated binding of those transcription factors to their motifs. These high chromatin accessibilities decreased as they differentiated into retinal ganglion cells and non-retinal ganglion cells. Although that study is excellent in providing comprehensive and in-depth insights, ours is unique in focusing on the Notch loci for clarifying the regulatory mechanisms in view of Notch signaling biology.
Finally, we note that further investigations, such as a large deletion of these regions, will be needed to evaluate the contribution of the identified chromatin remodeling to the differential expression of Notch receptors in RPC subsets.
Y. W.: Data Curation, Formal Analysis, Software, Visualization, Writing – Original Draft Preparation
S.K.: Writing – Original Draft Preparation
T.N.: Writing – Original Draft Preparation
S.T.: Supervision, Writing – Review & Editing
M.Y.: Conceptualization, Formal Analysis, Funding Acquisition, Methodology, Project Administration, Software, Writing – Original Draft Preparation
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)