Hamrayev D, Karimov B, Esanov H et al. Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.12688/f1000research.178398.3)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
1Institute of Botany, Academy of Sciences of Uzbekistan, Tashkent, Uzbekistan 2Bukhara State University, Bukhara, Uzbekistan 3Navoi State University, Navoi, Uzbekistan 4Samarkand State Medical University, Samarkand, Uzbekistan 5Bukhara state medical institute named after Abu Ali ibn Sino, Bukhara, Uzbekistan 6Karshi State University, Qarshi, Kashkadarya Province, Uzbekistan 7Turan University, Karshi, Uzbekistan 8Jizzakh State Pedagogical University, Jizzakh, Uzbekistan 9Tashkent Botanical Garden under the Institute of Botany, Academy of Sciences, Tashkent, Uzbekistan
Diyorjon Hamrayev
Roles:
Writing – Original Draft Preparation
Bobur Karimov
Roles:
Writing – Original Draft Preparation,
Writing – Review & Editing
Husniddin Esanov
Roles:
Writing – Original Draft Preparation
Sodik Khuzhzhiev
Roles:
Writing – Original Draft Preparation
Maxzuna Nasretdinova
Roles:
Writing – Original Draft Preparation
Oysha Jabborova
Roles:
Writing – Original Draft Preparation
Oybek Omonov
Roles:
Writing – Original Draft Preparation
Mohistara Sharipova
Roles:
Writing – Original Draft Preparation
This article is included in the Plant Science gateway.
Abstract
Chloroplast genomes provide important insights into plant phylogeny, genome evolution, and molecular marker development. In this study, we sequenced, assembled, and analyzed the complete chloroplast genomes of two endemic species from Uzbekistan, Astragalus nuratensis and Oxytropis pseudorosea. Genome skimming produced high-quality paired-end reads, yielding mean chloroplast sequencing depths of 638.13× for Astragalus nuratensis and 1,725.34× for Oxytropis pseudorosea, respectively. The chloroplast genomes were 122,316 bp in A. nuratensis and 122,708 bp in O. pseudorosea. Both genomes encoded 110 unique genes, including 76 protein-coding genes, 30 transfer RNA genes, and 4 ribosomal RNA genes. Consistent with members of the inverted repeat–lacking clade of Fabaceae, both species lacked the typical inverted repeat regions, resulting in a single-copy genome structure. Phylogenetic analysis based on 119 complete chloroplast genomes resolved major lineages within Astragalus and related genera with strong support. Astragalus nuratensis was placed within the Phaca clade, while Oxytropis pseudorosea formed part of a distinct Oxytropis lineage. These results provide new genomic resources for understanding evolutionary relationships and plastome evolution in Central Asian legumes.
Keywords
plastome; Fabaceae; IRLC; phylogeny; genome evolution; Central Asia
Corresponding author:
Ziyoviddin Yusupov
Competing interests:
No competing interests were disclosed.
Grant information:
This work was supported by the project “Digital Nature: Development of a digital platform for the flora of Central Uzbekistan” (2025–2029), implemented by the Institute of Botany, Academy of Sciences of the Republic of Uzbekistan. Additional support was provided by the project “Assessing climate change adaptation in endangered plants of Uzbekistan: A DNA barcoding approach” (AL-9224104464). The funders had no role in study design, data collection, analysis, decision to publish, or preparation of the manuscript.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
The manuscript has been updated to correct the italic formatting of gene names (rps16, rpl22, infA, accD, ycf4, clpP, atpF, and rpoC1) and the genus name Oxytropis in the Introduction, as suggested by the reviewer.
The manuscript has been updated to correct the italic formatting of gene names (rps16, rpl22, infA, accD, ycf4, clpP, atpF, and rpoC1) and the genus name Oxytropis in the Introduction, as suggested by the reviewer.
Chloroplast (cp) genomes of angiosperms have been widely used in studies of phylogeny, sequence variation, genome evolution, and the development of molecular markers (Dong et al., 2013). In most angiosperms, the cp genome exhibits a conserved quadripartite structure consisting of a large single-copy (LSC) region, a small single-copy (SSC) region, and two inverted repeat (IR) regions (Sugiura, 1992). Nevertheless, structural modifications, including gene rearrangements, inversions, expansion or loss of IR regions, gene loss, and pseudogene formation, have been documented in several lineages, particularly in Fabaceae (Cai et al., 2008; Son & Choi, 2022).
Fabaceae is one of the largest families of flowering plants and includes many species of considerable ecological and agricultural importance. According to recent classifications, the family is divided into six subfamilies: Caesalpinioideae, Cercidoideae, Detarioideae, Dialioideae, Duparquetioideae, and Faboideae (Papilionoideae) (Azani et al., 2017). Within Faboideae, the inverted repeat–lacking clade (IRLC) is characterized by the loss of one copy of the IR region (approximately 25 kb) in the chloroplast genome. This clade comprises about 52 genera and more than 4,000 species, and plastomes of IRLC taxa frequently show gene loss or pseudogenization (e.g., rps16, rpl22, infA, accD, and ycf4), intron loss (clpP, atpF, and rpoC1), inversions, and occasional transfer of genes to the nuclear genome (Son & Choi, 2022).
Within the IRLC, the genera Astragalus L. (3,095 species) and Oxytropis DC. (608 species) represent highly diverse and taxonomically complex groups (POWO, 2026). In recent years, complete chloroplast genome sequencing has been increasingly applied to resolve phylogenetic relationships and species boundaries within the genera Astragalus and Oxytropis. Several chloroplast genomes of species belonging to these genera have been published (Su et al., 2021; Li et al., 2025; Bobur et al., 2026). Despite these advances, chloroplast genome data remain limited for many narrowly distributed endemic species from Uzbekistan. In the present study, we sequenced, assembled, and analyzed the cp genomes of Oxytropis pseudorosea Filim. and Astragalus nuratensis Popov, two narrowly distributed endemic species from the Nuratau Mountains of Uzbekistan, Central Asia.
Methods
Leaf samples of Oxytropis pseudorosea and Astragalus nuratensis were collected in 2024 from the Nuratau Mountains, Uzbekistan, by Diyorjon Hamrayev. Astragalus nuratensis was identified by Beshko Natalya and its voucher specimen was deposited in the National Herbarium of Uzbekistan (TASH) under accession number TASH-2024-AN-001, whereas Oxytropis pseudorosea was identified by Doston Turdiyev and its voucher specimen was deposited in TASH under accession number TASH-2024-OP-002.
Genomic DNA was isolated from leaf tissue using the Tiangen DP305 Plant Genomic DNA Kit (Beijing, China). Libraries were prepared with the NEBNext® Ultra™ DNA Library Prep Kit for Illumina (NEB, USA; Cat. E7370L) following the manufacturer’s protocol, with index codes added during preparation. DNA was sonicated to ~350 bp and fragments were end-repaired, A-tailed, and ligated to Illumina adapters, followed by PCR amplification. PCR products were purified using AMPure XP beads (Beverly, USA). Library quality was assessed on an Agilent 5400 system, and concentrations were quantified by qPCR (1.5 nM). Qualified libraries were pooled and sequenced on Illumina platforms using the PE150 strategy at Novogene (Beijing, China).
Raw reads were assembled de novo using NOVOPlasty v4.3.5 (Dierckxsens et al., 2017), with the plastomes of Astragalus agrestis Douglas ex G. Don and Oxytropis neimonggolica C.W. Chang & Y.Z. Zhao serving as seed references. The assembly generated a single circular chloroplast genome for each species without structural ambiguities. To assess assembly accuracy and sequencing depth, clean paired-end reads were aligned to the assembled plastomes using BWA-MEM v0.7.17 (Li, 2013). Alignments were subsequently sorted and indexed with SAMtools v1.19.2 (Li et al., 2009). Genome annotation was conducted in Geneious v9.0.2 (Kearse et al., 2012) using closely related reference plastomes. Protein-coding genes, transfer RNAs, and ribosomal RNAs were annotated, and gene boundaries were manually curated to ensure accurate start/stop codons and intron–exon junctions. Furthermore, the cp map was generated using Chloroplot (https://irscope.shinyapps.io/chloroplot/) (Zheng et al., 2020).
For several taxa included in the phylogenetic analysis, complete chloroplast genome sequences were not available in GenBank. Only raw sequencing data (SRA accessions; SRR numbers listed in
Table 1) were available. Therefore, the chloroplast genomes of these taxa were assembled de novo using NOVOPlasty v4.3.5 (Dierckxsens et al., 2017). A total of 119 complete chloroplast genome sequences were included in the phylogenetic analysis. Of these, two plastomes were newly sequenced in this study, while the remaining 117 sequences were downloaded from the NCBI GenBank database (
Table 1). Complete chloroplast genome sequences were aligned using MAFFT (Katoh & Standley, 2013). The alignment was manually inspected and used to reconstruct phylogenetic relationships under the maximum likelihood criterion in IQ-TREE 2 (Minh et al., 2020). The best-fit substitution model was selected using ModelFinder (Kalyaanamoorthy et al., 2017), and branch support was assessed with 1,000 ultrafast bootstrap replicates (Hoang et al., 2018).
Table 1. List of taxa used in the phylogenetic analysis, including their respective GenBank accession numbers.
For taxa lacking published chloroplast genome accessions, plastomes were assembled in this study from raw reads downloaded from the NCBI Sequence Read Archive (SRR accessions provided).
Species
Accession
Species
Accession
Astragalus camptodontus
SRR12136586
A. aksuensis
SRR12136588
A. confertus
SRR12136589
A. massanderanus
SRR12136591
A. handelii
SRR12136594
A. zacharensis
SRR12136595
A. uliginosus
SRR12136597
A. koburensis
SRR12136600
A. beketowii
SRR12136611
A. jessenii
SRR12136613
A. tibetanus
SRR12136615
A. dendroides
SRR12136619
A. candissimus
SRR12136622
A. platyphyllus
SRR12136623
A. himalayanus
SRR12136624
A. roseus
SRR12136625
A. stalinskii
SRR12136626
A. orbicularifolius
SRR12136627
A. hamosus
SRR12136635
A. heinsensis
SRR12136637
A. hoantchy
SRR12136647
A. saccolacys
SRR12136648
A. neomondelphus
SRR12136653
A. schmalhausenii
SRR12136654
A. zingeri
SRR12136656
A. eximius
SRR12136657
A. xanthomeloides
SRR12136658
A. piletocladus
SRR12136659
A. parrowianus
SRR12136660
A. coluteocarpus
SRR12136661
A. thurberi
SRR12136664
A. americanus
SRR12136665
A. peduncularis
SRR12136666
A. dipelta
SRR12136669
A. acaulis
PV802397
A. austrotristis
NC_063479
A. adsurgens
NC_085710
A. angustidens
ON550413
A. agrestis
PP328800
A. alpinus
NC_063482
A. ampullarius
PV870668
A. arbuscula
ON550410
A. arpilobus
NC_063483
A. arrectus
NC_047381
A. bhotanensis
NC_063484
A. biristatus
MZ923743
A. calycosus var. calycosus
MZ725323
A. canadensis var. brevidens
PV870770
A. chinensis
NC_063486
A. clevelandii
ON550409
A. cognatus
MZ127832
A. chinense
ON550407
A. dilutus
PV870843
A. effusus
MZ901207
A. flexuosus var. flexuosus
ON550403
A. flexus
NC_083391
A. floridus
NC_058825
A. galactites
OY754317
A. glycyphyllos
MY746310
A. gummifer
NC_063487
A. gypsodes
LC764834
A. iranicus
NC_077545
A. juanquensis
PV798172
A. khasianus
ON550401
A. laksamanni
NC_085734
A. leucocephalus
ON550399
A. lithophilus
PV870772
A. lonchocarpus
LC764835
A. macropelmatus
OP723863
A. melilotoides var. tenuis
OP723862
A. membranaceus
PV761671
A. membranaceus var. membranaceus
KX255662
A. moseleios
LC764836
A. mongholicus
OR712437
A. mongholicus var. dahuricus
MW719856
A. mongholicus var. nakaianus
KR296789
A. muliensis
NC_083909
A. neglectus
NC_063488
A. nuratensis
PX928918
A. obscurus
NC_063489
A. odoratus
LC764837
A. oxylottis
NC_077543
A. pallasi
NC_077542
A. polysladus
PV910879
A. psilosepalus
NC_083910
A. purpurinus
OR652287
A. rumpens
PX711949
A. scaberrimus
MW654102
A. sereno var. sereno
MZ923754
A. sieversianus
NC_077540
A. sinicus
OM287552
A. stipulatus
OR491698
A. strictus
MT120746
A. toanus var. toanus
MZ923756
A. tongolensis
PX754582
A. tumabtsica
NC_083912
A. vulpinus
ON550388
A. wootonii var. wootonii
MZ923757
A. yunnanensis subsp. incanus
PV156653
A. zerabulaki
PX711948
Oxytropis chiliophylla
PV694277
O. filiformis
PV684033
O. latibracteata
PV694278
Oplonia microphylla
PV684029
O. neimongolica
PV684030
O. proboscidea
PV694280
O. pseudorosea
PX512474
O. sp. TT4869–1
PX652196
Tibetia liangshanensis
MF193597
Alhagi sparsifolia
MW349013
Caragana korshinskii
KX289923
Hedysarum taipeicum
MK426698
Caragana microphylla
KX289922
Corethrodendron multijugum
OP748243
Carmichaelia australis
PV870797
Phyllolobium chinense
MZ221114
Phyllolobium camptodontum
SRR12136586
Lessertia frutescens
MF286764
Sphaerophysa salsula
PV870675
Results
Genome skimming generated a total of 39,668,027 paired-end reads for A. nuratensis and 16,998,801 paired-end reads for O. pseudorosea. After quality filtering, 39,666,696 and 16,994,332 reads were retained as high-quality reads for A. nuratensis and O. pseudorosea, respectively. Mapping of the filtered reads to the assembled chloroplast reference genomes showed that 540,829 reads (1.36%) in A. nuratensis and 1,421,542 reads (8.36%) in O. pseudorosea were successfully aligned. Properly paired reads accounted for 1.33% and 8.27% of the total reads, respectively. The chloroplast genome of Astragalus nuratensis was recovered with an average sequencing depth of 638.13×, with coverage ranging from 9× to 3273×. In comparison, the chloroplast genome of Oxytropis pseudorosea exhibited a substantially higher mean sequencing depth of 1,725.34×, with coverage ranging from 12× to 2146× (Figure 1).
Figure 1. Coverage depth across the chloroplast genomes of Astragalus nuratensis and Oxytropis pseudorosea.
The complete chloroplast genome of O. pseudorosea was 122,708 bp in length, whereas that of A. nuratensis was 122,316 bp. Both genomes encoded a total of 110 unique genes, including 77 protein-coding genes (CDS), 30 transfer RNA (tRNA) genes, and 4 ribosomal RNA (rRNA) genes (
Figure 2). In both Astragalus nuratensis and Oxytropis pseudorosea, the genes rpl16, rpl2, rpoC1, ndhA, ndhB, petB, petD, atpF, clpP, trnA-UGC, trnG-UCC, trnI-GAU, trnK-UUU, trnL-UAA, and trnV-UAC each contain one intron, while ycf3 contains two introns. In addition, rps12 is a trans-spliced gene with two introns (one cis-spliced and one trans-spliced). Consistent with other members of the inverted repeat–lacking clade (IRLC) of legumes, the typical inverted repeat regions were absent in both species, resulting in a single-copy chloroplast genome structure, a feature commonly reported in plastome studies of Astragalus and related taxa (Moghaddam et al., 2023; Ma et al., 2025; Li et al., 2025).
Figure 2. The circular maps of the chloroplast genomes of Oxytropis pseudorosea and Astragalus nuratensis were drawn using Chloroplot.
The innermost circle represents the lengths of chloroplast genomes. The second concentric ring illustrates nucleotide composition, where the orange segment indicates GC content and the yellow segment corresponds to AT content. The outermost circle displays the distribution of genes, which are color-coded according to their functional classification. The transcriptional orientation is indicated such that genes on the inner circle are transcribed clockwise, whereas those on the outer circle are transcribed anticlockwise.
Maximum likelihood analysis based on complete chloroplast genome sequences resolved the sampled taxa into several well-supported clades corresponding to recognized infrageneric groups within Astragalus and related genera (
Figure 3). Most backbone nodes received strong bootstrap support (BS ≥ 95), indicating a stable phylogenetic structure. The overall topology was consistent with previous phylogenomic studies of Astragalus, which also recovered major lineages with strong support using plastome and target-enrichment data (Su et al., 2021; Buono et al., 2025).
Figure 3. Maximum likelihood phylogenetic tree inferred from complete chloroplast genome sequences showing the relationships of Astragalus nuratensis, Oxytropis pseudorosea, and other members of the inverted repeat–lacking clade (IRLC).
Species of Astragalus were distributed among the major lineages corresponding to the Hypoglottis, Neo-Astragalus, Diholcos, Astracantha, Contortuplicata, Hamosa, Trimeniaeus, and Phaca clades. Among these, the Phaca clade represented one of the largest lineages and included A. nuratensis, which grouped with other members of this clade with strong bootstrap support. Species of Oxytropis, including O. pseudorosea, formed a distinct and well-supported lineage corresponding to the Oxytropis + Coluteoid clade, clearly separated from the main Astragalus lineages, in agreement with previous plastome-based phylogenies (Buono et al., 2025).
In the phylogenetic tree, A. zerabulaki, another endemic species, was resolved within the Hypoglottis clade and formed a well-supported sister relationship with A. rumpens, indicating close evolutionary relationships among members of this lineage.
Azani N, Babineau M, Bailey CD, et al.:
A New Subfamily Classification of the Leguminosae Based on a Taxonomically Comprehensive Phylogeny: The Legume Phylogeny Working Group (LPWG).
Taxon.
2017; 66: 44–77. Publisher Full Text
Bobur K, Ziyoviddin Y, Ergashali R, et al.:
Chloroplast genome evidence in assessing the taxonomic status of Astragalus zerabulaki Popov (Fabaceae), an endemic of Uzbekistan.
Phytotaxa.
2026; 744(3): 249–260. Publisher Full Text
Buono D, Kadereit G, Liston A, et al.:
Building a robust backbone for Astragalus using a clade-specific target enrichment bait set.
Am. J. Bot.
2025; 112(8): e70084. Publisher Full Text
Cai Z, Guisinger M, Kim H-G, et al.:
Extensive Reorganization of the Plastid Genome of Trifolium Subterraneum (Fabaceae) Is Associated with Numerous Repeated Sequences and Novel DNA Insertions.
J. Mol. Evol.
2008; 67: 696–704. Publisher Full Text
Dong W, Xu C, Cheng T, et al.:
Sequencing angiosperm plastid genomes made easy: a complete set of universal primers and a case study on the phylogeny of Saxifragales.
Genome Biol. Evol.
2013; 5(5): 989–997. PubMed Abstract
| Publisher Full Text
| Free Full Text
Dierckxsens N, Mardulyn P, Smits G:
NOVOPlasty: de novo assembly of organelle genomes from whole genome data.
Nucleic Acids Res.
2017; 45(4): e18.
Hoang DT, Chernomor O, von Haeseler A , et al.:
UFBoot2: Improving the ultrafast bootstrap approximation.
Mol. Biol. Evol.
2018; 35(2): 518–522. Publisher Full Text
Kalyaanamoorthy S, Minh BQ, Wong TKF, et al.:
ModelFinder: Fast model selection for accurate phylogenetic estimates.
Nat. Methods.
2017; 14: 587–589. Publisher Full Text
Karimov B:
Raw sequencing data of Astragalus nuratensis and Oxytropis pseudorosea. [Dataset].
NCBI BioProject. PRJNA1425239.
2026a.
Reference Source
Karimov B:
Raw sequencing reads of Astragalus nuratensis and Oxytropis pseudorosea. [Dataset].
NCBI Sequence Read Archive (SRA). SRR37271903, SRR37271902.
2026b.
Reference Source
Karimov B:
Chloroplast genomes of Astragalus nuratensis and Oxytropis pseudorosea. [Dataset].
GenBank. PX928918, PX512474.
2026c.
Reference Source
Karimov B, Tojibaev SK, Azimova D, et al.:
Comparative analysis of complete chloroplast genomes of Cousinia (Asteraceae) species.
Front. Plant Sci.
2025; 16: 1522950. Publisher Full Text
Katoh K, Standley DM:
MAFFT multiple sequence alignment software version 7: Improvements in performance and usability.
Mol. Biol. Evol.
2013; 30(4): 772–780. Publisher Full Text
Kearse M, Moir R, Wilson A, et al.:
Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data.
Bioinformatics.
2012; 28(12): 1647–1649. Publisher Full Text
Li H:
Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM.
arXiv preprint arxiv:1303.3997.
2013. Publisher Full Text
Li H, Handsaker B, Wysoker A, et al.:
The sequence alignment/map format and SAMtools.
Bioinformatics.
2009; 25(16): 2078–2079. Publisher Full Text
Li QQ, Niu Y, Zhang ZP, et al.:
Comparative chloroplast genome analyses of Oxytropis DC. species: new insights into genome evolution and phylogenomic implications.
Front. Plant Sci.
2025; 16: 1645582. Publisher Full Text
Ma HT, Chen QY, Rao JH, et al.:
A Comprehensive Comparative Analysis and Phylogenetic Investigation of the Chloroplast Genome Sequences in Four Astragalus Species.
Curr. Issues Mol. Biol.
2025; 47(12): 978. Publisher Full Text
Minh BQ, Schmidt HA, Chernomor O, et al.:
IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era.
Mol. Biol. Evol.
2020; 37(5): 1530–1534. Publisher Full Text
Moghaddam M, Wojciechowski MF, Kazempour-Osaloo S:
Characterization and comparative analysis of the complete plastid genomes of four Astragalus species.
PLoS One.
2023; 18(5): e0286083. Publisher Full Text
POWO:
Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew.2026. Published on the Internet. Retrieved 17 February 2026.
Reference Source
Son O, Choi KS:
Characterization of the chloroplast genome structure of Gueldenstaedtia verna (Papilionoideae) and comparative analyses among IRLC species.
Forests.
2022; 13(11): 1942. Publisher Full Text
Su C, Duan L, Liu P, et al.:
Chloroplast phylogenomics and character evolution of eastern Asian Astragalus (Leguminosae): Tackling the phylogenetic structure of the largest genus of flowering plants in Asia.
Mol. Phylogenet. Evol.
2021; 156: 107025. Publisher Full Text
Sugiura M:
The chloroplast genome.
Plant Mol. Biol.
1992; 19: 149–168. Publisher Full Text
Zheng S, Poczai P, Hyvönen J, et al.:
Chloroplot: an online program for the versatile plotting of organelle genomes.
Front. Genet.
2020; 11: 576124. Publisher Full Text
1
Institute of Botany, Academy of Sciences of Uzbekistan, Tashkent, Uzbekistan 2
Bukhara State University, Bukhara, Uzbekistan 3
Navoi State University, Navoi, Uzbekistan 4
Samarkand State Medical University, Samarkand, Uzbekistan 5
Bukhara state medical institute named after Abu Ali ibn Sino, Bukhara, Uzbekistan 6
Karshi State University, Qarshi, Kashkadarya Province, Uzbekistan 7
Turan University, Karshi, Uzbekistan 8
Jizzakh State Pedagogical University, Jizzakh, Uzbekistan 9
Tashkent Botanical Garden under the Institute of Botany, Academy of Sciences, Tashkent, Uzbekistan
Diyorjon Hamrayev
Roles:
Writing – Original Draft Preparation
Bobur Karimov
Roles:
Writing – Original Draft Preparation,
Writing – Review & Editing
Husniddin Esanov
Roles:
Writing – Original Draft Preparation
Sodik Khuzhzhiev
Roles:
Writing – Original Draft Preparation
Maxzuna Nasretdinova
Roles:
Writing – Original Draft Preparation
Oysha Jabborova
Roles:
Writing – Original Draft Preparation
Oybek Omonov
Roles:
Writing – Original Draft Preparation
Mohistara Sharipova
Roles:
Writing – Original Draft Preparation
This work was supported by the project “Digital Nature: Development of a digital platform for the flora of Central Uzbekistan” (2025–2029), implemented by the Institute of Botany, Academy of Sciences of the Republic of Uzbekistan. Additional support was provided by the project “Assessing climate change adaptation in endangered plants of Uzbekistan: A DNA barcoding approach” (AL-9224104464). The funders had no role in study design, data collection, analysis, decision to publish, or preparation of the manuscript.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Hamrayev D, Karimov B, Esanov H et al. Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.12688/f1000research.178398.3)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
Do HDK. Reviewer Report For: Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.5256/f1000research.202456.r489947)
Dear Authors,
In the Introduction section, there are two sentences.
"This clade comprises about 52 genera and more than 4,000 species, and plastomes of IRLC taxa frequently show gene loss or pseudogenization (e.g., rps16, rpl22, infA, accD, and
... Continue reading
Dear Authors,
In the Introduction section, there are two sentences.
"This clade comprises about 52 genera and more than 4,000 species, and plastomes of IRLC taxa frequently show gene loss or pseudogenization (e.g., rps16, rpl22, infA, accD, and ycf4), intron loss (clpP, atpF, and rpoC1), inversions, and occasional transfer of genes to the nuclear genome (Son & Choi, 2022)."
"Within the IRLC, the genera Astragalus L. (3,095 species) and Oxytropis DC. (608 species) represent highly diverse and taxonomically complex groups (POWO, 2026)."
I am not sure the reason why the gene names and genus names were not italic in the above two sentences although the authors responsed that they fixed it.
Please check and revise.
Thank you.
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Plant Chloroplast genomes
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Do HDK. Reviewer Report For: Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.5256/f1000research.202456.r489947)
Do HDK. Reviewer Report For: Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.5256/f1000research.200633.r484854)
Dear Authors,
Thank you very much for revising the manuscript based on the reviewer's comments.
Most of my concerns were resolved.
However, there is still a minor issue about the italic font for gene names (rps16, rpl22,
... Continue reading
Dear Authors,
Thank you very much for revising the manuscript based on the reviewer's comments.
Most of my concerns were resolved.
However, there is still a minor issue about the italic font for gene names (rps16, rpl22, infA, accD, and ycf4; clpP, atpF, and rpoC1) and genus name (Oxytropis) in the Introduction.
Thank you.
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Plant Chloroplast genomes
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Do HDK. Reviewer Report For: Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.5256/f1000research.200633.r484854)
Bobur Karimov, Institute of Botany, Academy of Sciences of Uzbekistan, Tashkent, Uzbekistan
01 Jun 2026
Author Response
Dear Reviewer,
Thank you very much for your careful review and for your positive comments on our revised manuscript.
We appreciate your additional observation regarding the formatting issue. We
...
Continue readingDear Reviewer,
Thank you very much for your careful review and for your positive comments on our revised manuscript.
We appreciate your additional observation regarding the formatting issue. We have carefully checked the manuscript and corrected the italic formatting for all gene names (rps16, rpl22, infA, accD, ycf4, clpP, atpF, and rpoC1) as well as the genus name Oxytropis in the Introduction.
These corrections have been incorporated into the revised manuscript.
Thank you again for your valuable comments and suggestions.
Sincerely,
The Authors
Dear Reviewer,
Thank you very much for your careful review and for your positive comments on our revised manuscript.
We appreciate your additional observation regarding the formatting issue. We have carefully checked the manuscript and corrected the italic formatting for all gene names (rps16, rpl22, infA, accD, ycf4, clpP, atpF, and rpoC1) as well as the genus name Oxytropis in the Introduction.
These corrections have been incorporated into the revised manuscript.
Thank you again for your valuable comments and suggestions.
Sincerely,
The Authors
Competing Interests:No competing interests were disclosed.Close
Bobur Karimov, Institute of Botany, Academy of Sciences of Uzbekistan, Tashkent, Uzbekistan
01 Jun 2026
Author Response
Dear Reviewer,
Thank you very much for your careful review and for your positive comments on our revised manuscript.
We appreciate your additional observation regarding the formatting issue. We
...
Continue readingDear Reviewer,
Thank you very much for your careful review and for your positive comments on our revised manuscript.
We appreciate your additional observation regarding the formatting issue. We have carefully checked the manuscript and corrected the italic formatting for all gene names (rps16, rpl22, infA, accD, ycf4, clpP, atpF, and rpoC1) as well as the genus name Oxytropis in the Introduction.
These corrections have been incorporated into the revised manuscript.
Thank you again for your valuable comments and suggestions.
Sincerely,
The Authors
Dear Reviewer,
Thank you very much for your careful review and for your positive comments on our revised manuscript.
We appreciate your additional observation regarding the formatting issue. We have carefully checked the manuscript and corrected the italic formatting for all gene names (rps16, rpl22, infA, accD, ycf4, clpP, atpF, and rpoC1) as well as the genus name Oxytropis in the Introduction.
These corrections have been incorporated into the revised manuscript.
Thank you again for your valuable comments and suggestions.
Sincerely,
The Authors
Competing Interests:No competing interests were disclosed.Close
Fukasawa Y. Reviewer Report For: Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.5256/f1000research.200633.r484855)
Thank you for addressing the comments. The revised manuscript has been substantially improved. I
... Continue reading
Dear Authors,
Thank you for addressing the comments. The revised manuscript has been substantially improved. I am satisfied that the concerns raised in my initial review have been adequately addressed and am happy to approve this article.
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Plant genomics and bioinformatics
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Fukasawa Y. Reviewer Report For: Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.5256/f1000research.200633.r484855)
Fukasawa Y. Reviewer Report For: Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.5256/f1000research.196779.r473037)
Thank you for reporting the complete chloroplast genomes of Astragalus nuratensis and Oxytropis pseudorosea. The addition of new plastome resources for Astragalus and Oxytropis—two of the largest and most taxonomically challenging genera in the inverted
... Continue reading
Dear Authors,
Thank you for reporting the complete chloroplast genomes of Astragalus nuratensis and Oxytropis pseudorosea. The addition of new plastome resources for Astragalus and Oxytropis—two of the largest and most taxonomically challenging genera in the inverted repeat–lacking clade (IRLC)—is a welcome contribution, particularly given the ongoing interest in plastome evolution within this clade. Below I offer several suggestions that I believe would strengthen the manuscript.
1. Citation for the conserved quadripartite structure of chloroplast genomes
In the Introduction, the conserved quadripartite structure of angiosperm chloroplast genomes is cited as (Karimov et al. 2025). However, this is a well-established and broadly recognized feature of plastome architecture that has been described in numerous foundational works. The cited reference reports chloroplast genomes of Cousinia (Asteraceae) species and, while a valuable study in its own right, does not serve as a primary or representative source for this general statement. I would suggest replacing it with one or more widely cited references that originally established this structural model.
2. Chloroplast genome map of Oxytropis pseudorosea
Figure 1 presents the chloroplast genome map of A. nuratensis only. Since the manuscript reports two newly sequenced plastomes, I would recommend including a corresponding genome map of O. pseudorosea as well, either as an additional panel in Figure 1 or as a separate figure. This would provide a more balanced presentation of the two genomes.
3. Discrepancy in gene counts between Figure 1 and the text
The main text states that both genomes encode 76 protein-coding genes, 30 tRNA genes, and 4 rRNA genes (110 unique genes total). However, the annotation within Figure 1 indicates "77 genes; 4 rRNAs; 30 tRNAs" for A. nuratensis. Please verify and reconcile these numbers to ensure consistency between the figure and the text.
4. Assembly validation metrics
The authors report mapping clean reads back to the assembled plastomes using BWA-MEM, which is an appropriate approach. However, the current manuscript reports only the mean sequencing depth. To further support the completeness and accuracy of the assemblies, I would suggest briefly reporting additional validation metrics, such as: (i) whether any gaps or regions of zero coverage were observed, (ii) the minimum and maximum coverage across the genome, and/or (iii) a coverage depth plot provided as supplementary material. These additions would allow readers to better assess the quality of the assembled sequences.
5. Reporting gene loss, pseudogenization, and intron loss in the IRLC context
The Introduction appropriately highlights that IRLC plastomes frequently exhibit gene loss or pseudogenization (e.g., rps16, rpl22, infA, accD, ycf4) and intron loss (e.g., in clpP, atpF, rpoC1). However, the Results section does not describe which of these events, if any, were observed in the two newly sequenced plastomes. Even a brief statement confirming or contrasting the gene/intron content of A. nuratensis and O. pseudorosea with that of other IRLC members would add informative context to the genome descriptions and enhance the value of this report as a genomic resource.
Summary
This manuscript provides new plastome data for two IRLC species from Central Asia and meets the essential criteria for a Genome Note. The assembly and analytical approaches are technically appropriate.
Are the rationale for sequencing the genome and the species significance clearly described?
Yes
Are the protocols appropriate and is the work technically sound?
Yes
Are sufficient details of the sequencing and extraction, software used, and materials provided to allow replication by others?
Partly
Are the datasets clearly presented in a usable and accessible format, and the assembly and annotation available in an appropriate subject-specific repository?
Yes
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Plant genomics and bioinformatics
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Fukasawa Y. Reviewer Report For: Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.5256/f1000research.196779.r473037)
Bobur Karimov, Institute of Botany, Academy of Sciences of Uzbekistan, Tashkent, Uzbekistan
14 May 2026
Author Response
Reviewer 2
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the constructive comments, which have helped improve the clarity and quality of the study.
...
Continue readingReviewer 2
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the constructive comments, which have helped improve the clarity and quality of the study. Below we provide point-by-point responses.
1. Citation for the conserved quadripartite structure of chloroplast genomes
In the Introduction, the conserved quadripartite structure of angiosperm chloroplast genomes is cited as (Karimov et al. 2025). However, this is a well-established and broadly recognized feature of plastome architecture that has been described in numerous foundational works. The cited reference reports chloroplast genomes of Cousinia (Asteraceae) species and, while a valuable study in its own right, does not serve as a primary or representative source for this general statement. I would suggest replacing it with one or more widely cited references that originally established this structural model. Response: A more appropriate and widely accepted reference has been added to support the conserved quadripartite structure of angiosperm chloroplast genomes.
2. Chloroplast genome map of Oxytropis pseudorosea
Figure 1 presents the chloroplast genome map of A. nuratensis only. Since the manuscript reports two newly sequenced plastomes, I would recommend including a corresponding genome map of O. pseudorosea as well, either as an additional panel in Figure 1 or as a separate figure. This would provide a more balanced presentation of the two genomes. Response: The chloroplast genome map of Oxytropis pseudorosea has been added accordingly to provide balanced visualization of both plastomes.
3. Discrepancy in gene counts between Figure 1 and the text
The main text states that both genomes encode 76 protein-coding genes, 30 tRNA genes, and 4 rRNA genes (110 unique genes total). However, the annotation within Figure 1 indicates "77 genes; 4 rRNAs; 30 tRNAs" for A. nuratensis. Please verify and reconcile these numbers to ensure consistency between the figure and the text. Response: The gene annotation has been carefully checked, and inconsistencies between the figure and the text have been corrected to ensure full consistency.
4. Assembly validation metrics
The authors report mapping clean reads back to the assembled plastomes using BWA-MEM, which is an appropriate approach. However, the current manuscript reports only the mean sequencing depth. To further support the completeness and accuracy of the assemblies, I would suggest briefly reporting additional validation metrics, such as: (i) whether any gaps or regions of zero coverage were observed, (ii) the minimum and maximum coverage across the genome, and/or (iii) a coverage depth plot provided as supplementary material. These additions would allow readers to better assess the quality of the assembled sequences. Response: Additional assembly validation metrics have been included, including minimum and maximum coverage values, assessment of zero-coverage regions, and a coverage depth plot provided as Figure 1.
5. Reporting gene loss, pseudogenization, and intron loss in the IRLC context
The Introduction appropriately highlights that IRLC plastomes frequently exhibit gene loss or pseudogenization (e.g., rps16, rpl22, infA, accD, ycf4) and intron loss (e.g., in clpP, atpF, rpoC1). However, the Results section does not describe which of these events, if any, were observed in the two newly sequenced plastomes. Even a brief statement confirming or contrasting the gene/intron content of A. nuratensis and O. pseudorosea with that of other IRLC members would add informative context to the genome descriptions and enhance the value of this report as a genomic resource. Response: A brief description of gene content and intron structure has been added to the Results comparison with typical IRLC characteristics.
Reviewer 2
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the constructive comments, which have helped improve the clarity and quality of the study. Below we provide point-by-point responses.
1. Citation for the conserved quadripartite structure of chloroplast genomes
In the Introduction, the conserved quadripartite structure of angiosperm chloroplast genomes is cited as (Karimov et al. 2025). However, this is a well-established and broadly recognized feature of plastome architecture that has been described in numerous foundational works. The cited reference reports chloroplast genomes of Cousinia (Asteraceae) species and, while a valuable study in its own right, does not serve as a primary or representative source for this general statement. I would suggest replacing it with one or more widely cited references that originally established this structural model. Response: A more appropriate and widely accepted reference has been added to support the conserved quadripartite structure of angiosperm chloroplast genomes.
2. Chloroplast genome map of Oxytropis pseudorosea
Figure 1 presents the chloroplast genome map of A. nuratensis only. Since the manuscript reports two newly sequenced plastomes, I would recommend including a corresponding genome map of O. pseudorosea as well, either as an additional panel in Figure 1 or as a separate figure. This would provide a more balanced presentation of the two genomes. Response: The chloroplast genome map of Oxytropis pseudorosea has been added accordingly to provide balanced visualization of both plastomes.
3. Discrepancy in gene counts between Figure 1 and the text
The main text states that both genomes encode 76 protein-coding genes, 30 tRNA genes, and 4 rRNA genes (110 unique genes total). However, the annotation within Figure 1 indicates "77 genes; 4 rRNAs; 30 tRNAs" for A. nuratensis. Please verify and reconcile these numbers to ensure consistency between the figure and the text. Response: The gene annotation has been carefully checked, and inconsistencies between the figure and the text have been corrected to ensure full consistency.
4. Assembly validation metrics
The authors report mapping clean reads back to the assembled plastomes using BWA-MEM, which is an appropriate approach. However, the current manuscript reports only the mean sequencing depth. To further support the completeness and accuracy of the assemblies, I would suggest briefly reporting additional validation metrics, such as: (i) whether any gaps or regions of zero coverage were observed, (ii) the minimum and maximum coverage across the genome, and/or (iii) a coverage depth plot provided as supplementary material. These additions would allow readers to better assess the quality of the assembled sequences. Response: Additional assembly validation metrics have been included, including minimum and maximum coverage values, assessment of zero-coverage regions, and a coverage depth plot provided as Figure 1.
5. Reporting gene loss, pseudogenization, and intron loss in the IRLC context
The Introduction appropriately highlights that IRLC plastomes frequently exhibit gene loss or pseudogenization (e.g., rps16, rpl22, infA, accD, ycf4) and intron loss (e.g., in clpP, atpF, rpoC1). However, the Results section does not describe which of these events, if any, were observed in the two newly sequenced plastomes. Even a brief statement confirming or contrasting the gene/intron content of A. nuratensis and O. pseudorosea with that of other IRLC members would add informative context to the genome descriptions and enhance the value of this report as a genomic resource. Response: A brief description of gene content and intron structure has been added to the Results comparison with typical IRLC characteristics.
Competing Interests:No competing interests were disclosed.Close
Bobur Karimov, Institute of Botany, Academy of Sciences of Uzbekistan, Tashkent, Uzbekistan
14 May 2026
Author Response
Reviewer 2
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the constructive comments, which have helped improve the clarity and quality of the study.
...
Continue readingReviewer 2
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the constructive comments, which have helped improve the clarity and quality of the study. Below we provide point-by-point responses.
1. Citation for the conserved quadripartite structure of chloroplast genomes
In the Introduction, the conserved quadripartite structure of angiosperm chloroplast genomes is cited as (Karimov et al. 2025). However, this is a well-established and broadly recognized feature of plastome architecture that has been described in numerous foundational works. The cited reference reports chloroplast genomes of Cousinia (Asteraceae) species and, while a valuable study in its own right, does not serve as a primary or representative source for this general statement. I would suggest replacing it with one or more widely cited references that originally established this structural model. Response: A more appropriate and widely accepted reference has been added to support the conserved quadripartite structure of angiosperm chloroplast genomes.
2. Chloroplast genome map of Oxytropis pseudorosea
Figure 1 presents the chloroplast genome map of A. nuratensis only. Since the manuscript reports two newly sequenced plastomes, I would recommend including a corresponding genome map of O. pseudorosea as well, either as an additional panel in Figure 1 or as a separate figure. This would provide a more balanced presentation of the two genomes. Response: The chloroplast genome map of Oxytropis pseudorosea has been added accordingly to provide balanced visualization of both plastomes.
3. Discrepancy in gene counts between Figure 1 and the text
The main text states that both genomes encode 76 protein-coding genes, 30 tRNA genes, and 4 rRNA genes (110 unique genes total). However, the annotation within Figure 1 indicates "77 genes; 4 rRNAs; 30 tRNAs" for A. nuratensis. Please verify and reconcile these numbers to ensure consistency between the figure and the text. Response: The gene annotation has been carefully checked, and inconsistencies between the figure and the text have been corrected to ensure full consistency.
4. Assembly validation metrics
The authors report mapping clean reads back to the assembled plastomes using BWA-MEM, which is an appropriate approach. However, the current manuscript reports only the mean sequencing depth. To further support the completeness and accuracy of the assemblies, I would suggest briefly reporting additional validation metrics, such as: (i) whether any gaps or regions of zero coverage were observed, (ii) the minimum and maximum coverage across the genome, and/or (iii) a coverage depth plot provided as supplementary material. These additions would allow readers to better assess the quality of the assembled sequences. Response: Additional assembly validation metrics have been included, including minimum and maximum coverage values, assessment of zero-coverage regions, and a coverage depth plot provided as Figure 1.
5. Reporting gene loss, pseudogenization, and intron loss in the IRLC context
The Introduction appropriately highlights that IRLC plastomes frequently exhibit gene loss or pseudogenization (e.g., rps16, rpl22, infA, accD, ycf4) and intron loss (e.g., in clpP, atpF, rpoC1). However, the Results section does not describe which of these events, if any, were observed in the two newly sequenced plastomes. Even a brief statement confirming or contrasting the gene/intron content of A. nuratensis and O. pseudorosea with that of other IRLC members would add informative context to the genome descriptions and enhance the value of this report as a genomic resource. Response: A brief description of gene content and intron structure has been added to the Results comparison with typical IRLC characteristics.
Reviewer 2
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the constructive comments, which have helped improve the clarity and quality of the study. Below we provide point-by-point responses.
1. Citation for the conserved quadripartite structure of chloroplast genomes
In the Introduction, the conserved quadripartite structure of angiosperm chloroplast genomes is cited as (Karimov et al. 2025). However, this is a well-established and broadly recognized feature of plastome architecture that has been described in numerous foundational works. The cited reference reports chloroplast genomes of Cousinia (Asteraceae) species and, while a valuable study in its own right, does not serve as a primary or representative source for this general statement. I would suggest replacing it with one or more widely cited references that originally established this structural model. Response: A more appropriate and widely accepted reference has been added to support the conserved quadripartite structure of angiosperm chloroplast genomes.
2. Chloroplast genome map of Oxytropis pseudorosea
Figure 1 presents the chloroplast genome map of A. nuratensis only. Since the manuscript reports two newly sequenced plastomes, I would recommend including a corresponding genome map of O. pseudorosea as well, either as an additional panel in Figure 1 or as a separate figure. This would provide a more balanced presentation of the two genomes. Response: The chloroplast genome map of Oxytropis pseudorosea has been added accordingly to provide balanced visualization of both plastomes.
3. Discrepancy in gene counts between Figure 1 and the text
The main text states that both genomes encode 76 protein-coding genes, 30 tRNA genes, and 4 rRNA genes (110 unique genes total). However, the annotation within Figure 1 indicates "77 genes; 4 rRNAs; 30 tRNAs" for A. nuratensis. Please verify and reconcile these numbers to ensure consistency between the figure and the text. Response: The gene annotation has been carefully checked, and inconsistencies between the figure and the text have been corrected to ensure full consistency.
4. Assembly validation metrics
The authors report mapping clean reads back to the assembled plastomes using BWA-MEM, which is an appropriate approach. However, the current manuscript reports only the mean sequencing depth. To further support the completeness and accuracy of the assemblies, I would suggest briefly reporting additional validation metrics, such as: (i) whether any gaps or regions of zero coverage were observed, (ii) the minimum and maximum coverage across the genome, and/or (iii) a coverage depth plot provided as supplementary material. These additions would allow readers to better assess the quality of the assembled sequences. Response: Additional assembly validation metrics have been included, including minimum and maximum coverage values, assessment of zero-coverage regions, and a coverage depth plot provided as Figure 1.
5. Reporting gene loss, pseudogenization, and intron loss in the IRLC context
The Introduction appropriately highlights that IRLC plastomes frequently exhibit gene loss or pseudogenization (e.g., rps16, rpl22, infA, accD, ycf4) and intron loss (e.g., in clpP, atpF, rpoC1). However, the Results section does not describe which of these events, if any, were observed in the two newly sequenced plastomes. Even a brief statement confirming or contrasting the gene/intron content of A. nuratensis and O. pseudorosea with that of other IRLC members would add informative context to the genome descriptions and enhance the value of this report as a genomic resource. Response: A brief description of gene content and intron structure has been added to the Results comparison with typical IRLC characteristics.
Competing Interests:No competing interests were disclosed.Close
Do HDK. Reviewer Report For: Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.5256/f1000research.196779.r471852)
Dear Authors,
Thank you very much for adding new genomic data about Astragalus and Oxytropis species. The content of the manuscript is suitable for Genome Note of F1000 Research.
Please check my comment below.
1/ Please
... Continue reading
Dear Authors,
Thank you very much for adding new genomic data about Astragalus and Oxytropis species. The content of the manuscript is suitable for Genome Note of F1000 Research.
Please check my comment below.
1/ Please update the role of authors because most authors are for writing the manuscript. Who did the sampling, experiment, and analyzing the data?
2/ "Chloroplast (cp) genomes of angiosperms have been widely used in studies of phylogeny, sequence variation, genome evolution, and the development of molecular markers." Please add references for this statement.
3/ "In most angiosperms, the cp genome exhibits a conserved quadripartite structure consisting of a large single-copy (LSC) region, a small single-copy (SSC) region, and two inverted repeat (IR) regions (Karimov et al., 2025)." the current reference is not suitable for the statement because Karimov et al 2025 only reported complete chloroplast genomes of six Cousinia (Asteraceae) species which are not representative for most angiosperms.
4/ Please use italic font for gene names in the Introduction part.
5/ It would be better if the authors add introduction about the published chloroplast genomes of Astragalus and Oxytropis, especially Astragalus zerabulaki which was mentioned at the end of the manuscript.
6/ "Voucher specimens were deposited in the National Herbarium of Uzbekistan (TASH) under the accession numbers TASH-2024-AN-001 and TASH-2024-OP-002."; "Genome skimming generated high-quality paired-end reads, enabling the recovery of complete plastomes with mean sequencing depths of 638× and 1,725×, respectively." please revise these sentences to clarify which feature is of Astragalus nuratensis and which information belongs to Oxytropis pseudorosea.
7/ "High-quality reads were assembled de novo using NOVOPlasty v4.3.5" how to get high-quality reads? Please add.
8/ "For several taxa included in the phylogenetic analysis, complete chloroplast genome sequences were not available in GenBank. Only raw sequencing data (SRA accessions; SRR numbers listed in Table 1) were available. Therefore, the chloroplast genomes of these taxa were assembled." How to complete chloroplast genomes from SRA data? Please add.
9/ Because the manuscript reports two chloroplast genome, it would be better if the authors add the chloroplast genome map of O. pseudorosea in Figure 1.
10/ "Table 1. List of taxa included in the phylogenetic analysis, accession numbers." Please revise Table 1 caption.
11/ Please correct accession number of O. pseudorosea which is not PV684035 from GenBank database in Table 1.
Thank you.
Are the rationale for sequencing the genome and the species significance clearly described?
Yes
Are the protocols appropriate and is the work technically sound?
Yes
Are sufficient details of the sequencing and extraction, software used, and materials provided to allow replication by others?
Partly
Are the datasets clearly presented in a usable and accessible format, and the assembly and annotation available in an appropriate subject-specific repository?
Yes
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Plant Chloroplast genomes
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Do HDK. Reviewer Report For: Complete chloroplast genomes of endemic Astragalus and Oxytropis species from Uzbekistan [version 3; peer review: 1 approved, 1 approved with reservations]. F1000Research 2026, 15:445 (https://doi.org/10.5256/f1000research.196779.r471852)
Bobur Karimov, Institute of Botany, Academy of Sciences of Uzbekistan, Tashkent, Uzbekistan
14 May 2026
Author Response
Reviewer 1
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the valuable comments, which helped improve the quality of the work. Below we provide
...
Continue readingReviewer 1
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the valuable comments, which helped improve the quality of the work. Below we provide point-by-point responses.
Comment 1/ Please update the role of authors because most authors are for writing the manuscript. Who did the sampling, experiment, and analyzing the data? Response: The system did not provide an option to edit and resubmit the updated version with revised author contributions.
Comment 2/ "Chloroplast (cp) genomes of angiosperms have been widely used in studies of phylogeny, sequence variation, genome evolution, and the development of molecular markers." Please add references for this statement. Response: Appropriate references have been added to support this statement in the revised manuscript.
Comment 3/ "In most angiosperms, the cp genome exhibits a conserved quadripartite structure consisting of a large single-copy (LSC) region, a small single-copy (SSC) region, and two inverted repeat (IR) regions (Karimov et al., 2025)." the current reference is not suitable for the statement because Karimov et al 2025 only reported complete chloroplast genomes of six Cousinia (Asteraceae) species which are not representative for most angiosperms. Response: A more appropriate and widely accepted reference has been added to support this statement.
Comment 4/ Please use italic font for gene names in the Introduction part. Response: Gene names in the Introduction have been corrected and formatted in italics.
Comment 5/ It would be better if the authors add introduction about the published chloroplast genomes of Astragalus and Oxytropis, especially Astragalus zerabulaki which was mentioned at the end of the manuscript. Response: A new introductory paragraph has been added covering published chloroplast genomes of Astragalus and Oxytropis
Comment 6/ "Voucher specimens were deposited in the National Herbarium of Uzbekistan (TASH) under the accession numbers TASH-2024-AN-001 and TASH-2024-OP-002."; "Genome skimming generated high-quality paired-end reads, enabling the recovery of complete plastomes with mean sequencing depths of 638× and 1,725×, respectively." please revise these sentences to clarify which feature is of Astragalus nuratensis and which information belongs to Oxytropis pseudorosea. Response: The sentences have been revised to clearly distinguish species-specific voucher information and sequencing depth values.
Comment 7/ "High-quality reads were assembled de novo using NOVOPlasty v4.3.5" how to get high-quality reads? Please add. Response: Raw sequencing reads were directly used, and NOVOPlasty performs internal quality filtering during the assembly process.
Comment 8/ "For several taxa included in the phylogenetic analysis, complete chloroplast genome sequences were not available in GenBank. Only raw sequencing data (SRA accessions; SRR numbers listed in Table 1) were available. Therefore, the chloroplast genomes of these taxa were assembled." How to complete chloroplast genomes from SRA data? Please add.from SRA to novoplasty Response: Raw sequencing data were downloaded from SRA and assembled into chloroplast genomes using NOVOPlasty following the same assembly workflow.
Comment 9/ Because the manuscript reports two chloroplast genome, it would be better if the authors add the chloroplast genome map of O. pseudorosea in Figure 1. Response: The chloroplast genome map of Oxytropis pseudorosea has been added to Figure 1.
Comment 10/ "Table 1. List of taxa included in the phylogenetic analysis, accession numbers." Please revise Table 1 caption. Response: The caption of Table 1 has been revised for clarity and consistency.
Comment 11/ Please correct accession number of O. pseudorosea which is not PV684035 from GenBank database in Table 1. Response: The accession number has been corrected according to the GenBank database.
Reviewer 1
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the valuable comments, which helped improve the quality of the work. Below we provide point-by-point responses.
Comment 1/ Please update the role of authors because most authors are for writing the manuscript. Who did the sampling, experiment, and analyzing the data? Response: The system did not provide an option to edit and resubmit the updated version with revised author contributions.
Comment 2/ "Chloroplast (cp) genomes of angiosperms have been widely used in studies of phylogeny, sequence variation, genome evolution, and the development of molecular markers." Please add references for this statement. Response: Appropriate references have been added to support this statement in the revised manuscript.
Comment 3/ "In most angiosperms, the cp genome exhibits a conserved quadripartite structure consisting of a large single-copy (LSC) region, a small single-copy (SSC) region, and two inverted repeat (IR) regions (Karimov et al., 2025)." the current reference is not suitable for the statement because Karimov et al 2025 only reported complete chloroplast genomes of six Cousinia (Asteraceae) species which are not representative for most angiosperms. Response: A more appropriate and widely accepted reference has been added to support this statement.
Comment 4/ Please use italic font for gene names in the Introduction part. Response: Gene names in the Introduction have been corrected and formatted in italics.
Comment 5/ It would be better if the authors add introduction about the published chloroplast genomes of Astragalus and Oxytropis, especially Astragalus zerabulaki which was mentioned at the end of the manuscript. Response: A new introductory paragraph has been added covering published chloroplast genomes of Astragalus and Oxytropis
Comment 6/ "Voucher specimens were deposited in the National Herbarium of Uzbekistan (TASH) under the accession numbers TASH-2024-AN-001 and TASH-2024-OP-002."; "Genome skimming generated high-quality paired-end reads, enabling the recovery of complete plastomes with mean sequencing depths of 638× and 1,725×, respectively." please revise these sentences to clarify which feature is of Astragalus nuratensis and which information belongs to Oxytropis pseudorosea. Response: The sentences have been revised to clearly distinguish species-specific voucher information and sequencing depth values.
Comment 7/ "High-quality reads were assembled de novo using NOVOPlasty v4.3.5" how to get high-quality reads? Please add. Response: Raw sequencing reads were directly used, and NOVOPlasty performs internal quality filtering during the assembly process.
Comment 8/ "For several taxa included in the phylogenetic analysis, complete chloroplast genome sequences were not available in GenBank. Only raw sequencing data (SRA accessions; SRR numbers listed in Table 1) were available. Therefore, the chloroplast genomes of these taxa were assembled." How to complete chloroplast genomes from SRA data? Please add.from SRA to novoplasty Response: Raw sequencing data were downloaded from SRA and assembled into chloroplast genomes using NOVOPlasty following the same assembly workflow.
Comment 9/ Because the manuscript reports two chloroplast genome, it would be better if the authors add the chloroplast genome map of O. pseudorosea in Figure 1. Response: The chloroplast genome map of Oxytropis pseudorosea has been added to Figure 1.
Comment 10/ "Table 1. List of taxa included in the phylogenetic analysis, accession numbers." Please revise Table 1 caption. Response: The caption of Table 1 has been revised for clarity and consistency.
Comment 11/ Please correct accession number of O. pseudorosea which is not PV684035 from GenBank database in Table 1. Response: The accession number has been corrected according to the GenBank database.
Competing Interests:No competing interests were disclosed.Close
Bobur Karimov, Institute of Botany, Academy of Sciences of Uzbekistan, Tashkent, Uzbekistan
14 May 2026
Author Response
Reviewer 1
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the valuable comments, which helped improve the quality of the work. Below we provide
...
Continue readingReviewer 1
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the valuable comments, which helped improve the quality of the work. Below we provide point-by-point responses.
Comment 1/ Please update the role of authors because most authors are for writing the manuscript. Who did the sampling, experiment, and analyzing the data? Response: The system did not provide an option to edit and resubmit the updated version with revised author contributions.
Comment 2/ "Chloroplast (cp) genomes of angiosperms have been widely used in studies of phylogeny, sequence variation, genome evolution, and the development of molecular markers." Please add references for this statement. Response: Appropriate references have been added to support this statement in the revised manuscript.
Comment 3/ "In most angiosperms, the cp genome exhibits a conserved quadripartite structure consisting of a large single-copy (LSC) region, a small single-copy (SSC) region, and two inverted repeat (IR) regions (Karimov et al., 2025)." the current reference is not suitable for the statement because Karimov et al 2025 only reported complete chloroplast genomes of six Cousinia (Asteraceae) species which are not representative for most angiosperms. Response: A more appropriate and widely accepted reference has been added to support this statement.
Comment 4/ Please use italic font for gene names in the Introduction part. Response: Gene names in the Introduction have been corrected and formatted in italics.
Comment 5/ It would be better if the authors add introduction about the published chloroplast genomes of Astragalus and Oxytropis, especially Astragalus zerabulaki which was mentioned at the end of the manuscript. Response: A new introductory paragraph has been added covering published chloroplast genomes of Astragalus and Oxytropis
Comment 6/ "Voucher specimens were deposited in the National Herbarium of Uzbekistan (TASH) under the accession numbers TASH-2024-AN-001 and TASH-2024-OP-002."; "Genome skimming generated high-quality paired-end reads, enabling the recovery of complete plastomes with mean sequencing depths of 638× and 1,725×, respectively." please revise these sentences to clarify which feature is of Astragalus nuratensis and which information belongs to Oxytropis pseudorosea. Response: The sentences have been revised to clearly distinguish species-specific voucher information and sequencing depth values.
Comment 7/ "High-quality reads were assembled de novo using NOVOPlasty v4.3.5" how to get high-quality reads? Please add. Response: Raw sequencing reads were directly used, and NOVOPlasty performs internal quality filtering during the assembly process.
Comment 8/ "For several taxa included in the phylogenetic analysis, complete chloroplast genome sequences were not available in GenBank. Only raw sequencing data (SRA accessions; SRR numbers listed in Table 1) were available. Therefore, the chloroplast genomes of these taxa were assembled." How to complete chloroplast genomes from SRA data? Please add.from SRA to novoplasty Response: Raw sequencing data were downloaded from SRA and assembled into chloroplast genomes using NOVOPlasty following the same assembly workflow.
Comment 9/ Because the manuscript reports two chloroplast genome, it would be better if the authors add the chloroplast genome map of O. pseudorosea in Figure 1. Response: The chloroplast genome map of Oxytropis pseudorosea has been added to Figure 1.
Comment 10/ "Table 1. List of taxa included in the phylogenetic analysis, accession numbers." Please revise Table 1 caption. Response: The caption of Table 1 has been revised for clarity and consistency.
Comment 11/ Please correct accession number of O. pseudorosea which is not PV684035 from GenBank database in Table 1. Response: The accession number has been corrected according to the GenBank database.
Reviewer 1
We sincerely thank the reviewer for the careful evaluation of our manuscript and for the valuable comments, which helped improve the quality of the work. Below we provide point-by-point responses.
Comment 1/ Please update the role of authors because most authors are for writing the manuscript. Who did the sampling, experiment, and analyzing the data? Response: The system did not provide an option to edit and resubmit the updated version with revised author contributions.
Comment 2/ "Chloroplast (cp) genomes of angiosperms have been widely used in studies of phylogeny, sequence variation, genome evolution, and the development of molecular markers." Please add references for this statement. Response: Appropriate references have been added to support this statement in the revised manuscript.
Comment 3/ "In most angiosperms, the cp genome exhibits a conserved quadripartite structure consisting of a large single-copy (LSC) region, a small single-copy (SSC) region, and two inverted repeat (IR) regions (Karimov et al., 2025)." the current reference is not suitable for the statement because Karimov et al 2025 only reported complete chloroplast genomes of six Cousinia (Asteraceae) species which are not representative for most angiosperms. Response: A more appropriate and widely accepted reference has been added to support this statement.
Comment 4/ Please use italic font for gene names in the Introduction part. Response: Gene names in the Introduction have been corrected and formatted in italics.
Comment 5/ It would be better if the authors add introduction about the published chloroplast genomes of Astragalus and Oxytropis, especially Astragalus zerabulaki which was mentioned at the end of the manuscript. Response: A new introductory paragraph has been added covering published chloroplast genomes of Astragalus and Oxytropis
Comment 6/ "Voucher specimens were deposited in the National Herbarium of Uzbekistan (TASH) under the accession numbers TASH-2024-AN-001 and TASH-2024-OP-002."; "Genome skimming generated high-quality paired-end reads, enabling the recovery of complete plastomes with mean sequencing depths of 638× and 1,725×, respectively." please revise these sentences to clarify which feature is of Astragalus nuratensis and which information belongs to Oxytropis pseudorosea. Response: The sentences have been revised to clearly distinguish species-specific voucher information and sequencing depth values.
Comment 7/ "High-quality reads were assembled de novo using NOVOPlasty v4.3.5" how to get high-quality reads? Please add. Response: Raw sequencing reads were directly used, and NOVOPlasty performs internal quality filtering during the assembly process.
Comment 8/ "For several taxa included in the phylogenetic analysis, complete chloroplast genome sequences were not available in GenBank. Only raw sequencing data (SRA accessions; SRR numbers listed in Table 1) were available. Therefore, the chloroplast genomes of these taxa were assembled." How to complete chloroplast genomes from SRA data? Please add.from SRA to novoplasty Response: Raw sequencing data were downloaded from SRA and assembled into chloroplast genomes using NOVOPlasty following the same assembly workflow.
Comment 9/ Because the manuscript reports two chloroplast genome, it would be better if the authors add the chloroplast genome map of O. pseudorosea in Figure 1. Response: The chloroplast genome map of Oxytropis pseudorosea has been added to Figure 1.
Comment 10/ "Table 1. List of taxa included in the phylogenetic analysis, accession numbers." Please revise Table 1 caption. Response: The caption of Table 1 has been revised for clarity and consistency.
Comment 11/ Please correct accession number of O. pseudorosea which is not PV684035 from GenBank database in Table 1. Response: The accession number has been corrected according to the GenBank database.
Competing Interests:No competing interests were disclosed.Close
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
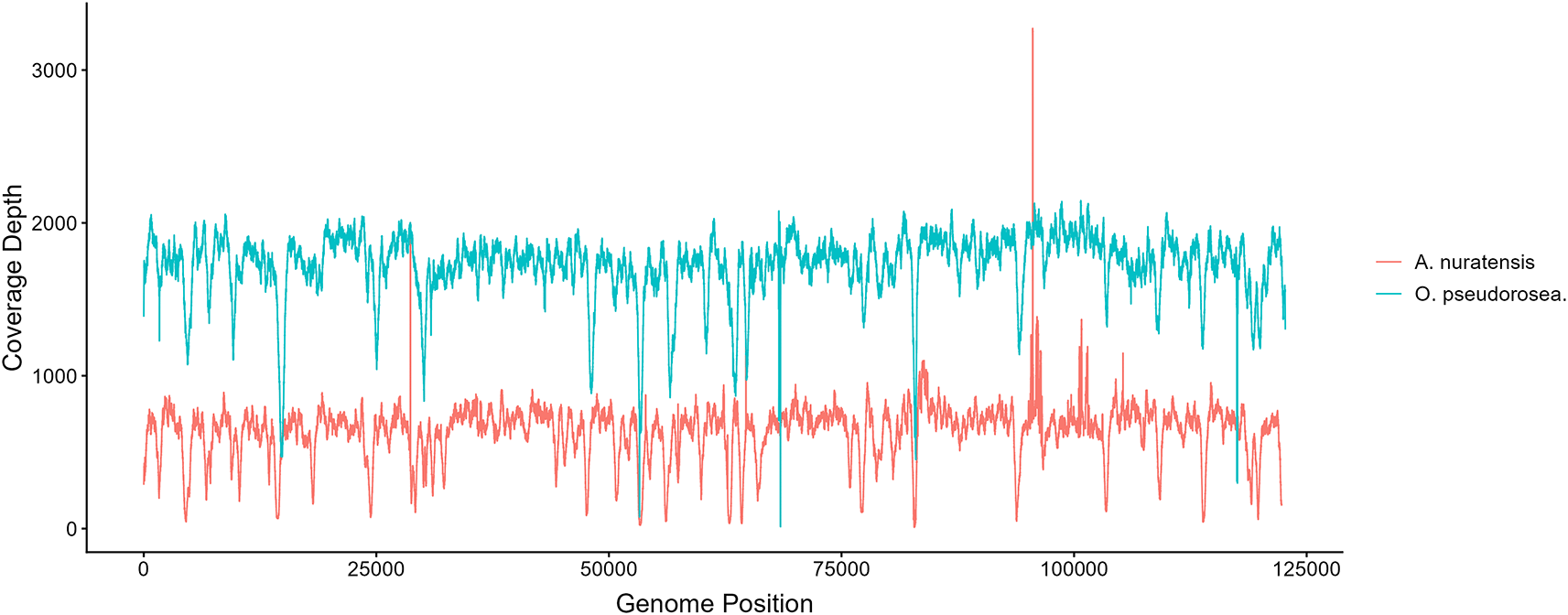
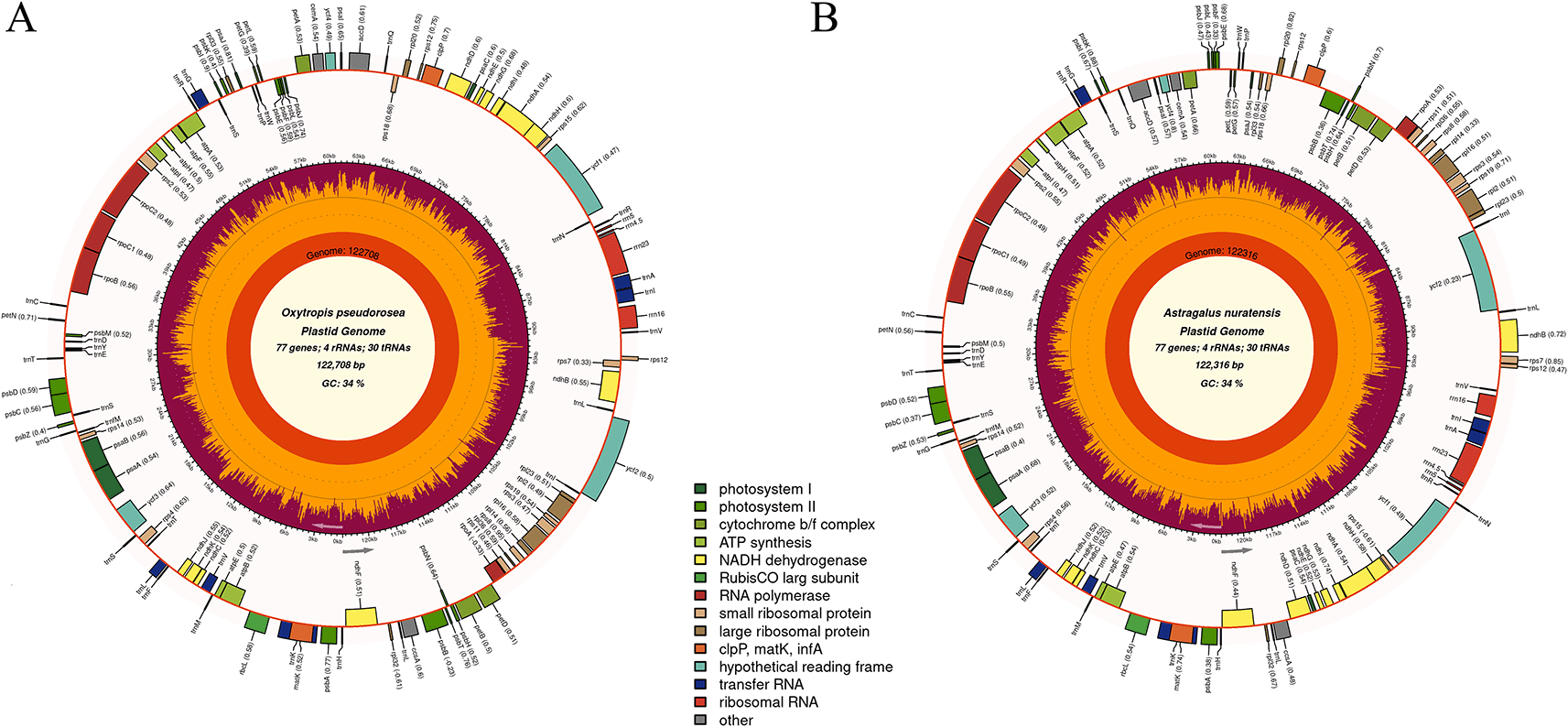
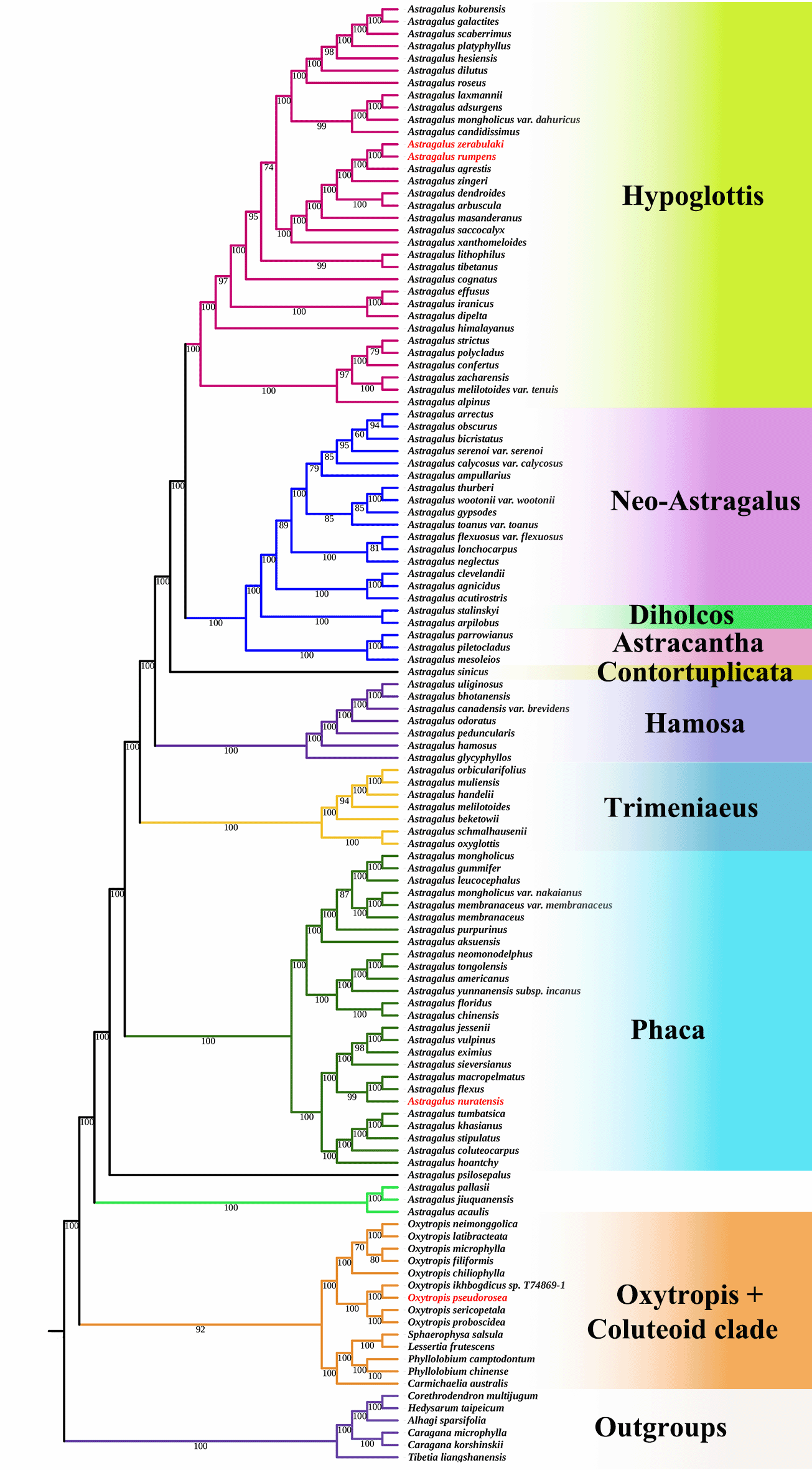
Comments on this article Comments (0)