Keywords
Plasmodium falciparum, Plasmodium vivax, malaria, animals, host reservoir, PCR.
Plasmodium falciparum, Plasmodium vivax, malaria, animals, host reservoir, PCR.
In this revised iteration of the article, significant enhancements have been implemented compared to its previously published counterpart. Primarily, meticulous attention has been devoted to refining the clarity and precision of language throughout the manuscript, addressing ambiguities and convoluted expressions to ensure a more coherent and accessible presentation of the research findings.
Moreover, the organization of content has been optimized to establish a more logical and seamless flow of information. Sections have been restructured, and transitions between different subsections have been strengthened to enhance the overall coherence of the narrative, facilitating a clearer understanding of the research methodology, results, and implications.
See the authors' detailed response to the review by Jaishree Raman
See the authors' detailed response to the review by Paul Cliff Simon Divis
See the authors' detailed response to the review by Alfred Amambua-Ngwa
Indonesia is striving toward becoming a malaria-free country through the Malaria Control Programme. Despite the implementation of several efforts, such as early detection, treatment, and mosquito vector eradication, the disease remains a persistent challenge. Factors including parasite resistance to antimalarial drugs, mosquito resistance to insecticides, inadequate health system performance, and host reservoir presence contribute to continuous malaria prevalence.
Plasmodium falciparum and P. vivax commonly infect humans, leading to high morbidity and mortality. Only humans were considered the primary host for Plasmodium species including P. falciparum, P. vivax, P. malariae, and P. ovale before molecular diagnostics development. However, studies over the past two decades reported that the parasites originated from animals. Specifically, P. falciparum was traced back to gorillas (Liu et al., 2010) and chimpanzees (Duval et al., 2010; Krief et al., 2010), while P. vivax was associated with African apes (Liu et al., 2014). Other sources stated were the origination of P. malariae from chimpanzees (Duval et al., 2010) and P. knowlesi from monkeys (Knowlesi, 1935; Zhang et al., 2016), while P. ovale found in humans and chimpanzees had genetic similarities (Duval et al., 2009). Factors such as habitat loss among primates and human encroachment into forested areas are believed to have facilitated the transmission of these parasites (Davidson et al., 2019). An investigation performed in South Kalimantan reported the contribution of forest workers to malaria incidence (Rahayu et al., 2016). Therefore, this study was conducted in Gaura village, West Sumba and Fakfak, West Papua, which contain a high percentage of forestry workers.
West Papua and East Nusa Tenggara are known as malaria-endemic provinces in Indonesia, with annual parasite incidence (API) of 7.04% and 31.29%, respectively in 2015 (Kemenkes, 2016), while the rates were 3.42% and 8.49% in 2018. Even though there was a decrease in API numbers, Fakfak, West Papua and East Nusa Tenggara, West Sumba, continued to report high API rates of 4.85% and 12.9%, respectively (Public Health Office of Fakfak and West Sumba, unpublished data). The trend of malaria cases varies, and despite reduced API rates recorded in several provinces, the two areas still experience a high incidence. Therefore, this study aimed to explore the presence of human Plasmodium among domestic animals, which might serve as a potential reservoir host in endemic areas. The exploration would provide insights into the reasons for high endemicity rates in the examined areas.
This study was conducted in October 2018 in Gaura village, West Sumba Regency, an area 29.96 km2 in size inhabited by 9,584 people, and Fakfak, West Papua Province, in August 2019 with an area of 11,036 km2 inhabited by 84,692 people (Figure 1). The residents’ main occupation is farming, while livestock such as goats, horses, cows, pigs, and buffalos are commonly found in their enclosures located around the owner’s residence. Furthermore, they also own pets such as dogs and cats. The average distance between enclosure and home in Fakfak was 225 meters while in Gaura village the distance was 0-10 meters.
Sampling was carried out by the veterinarian and staff from West Sumba and Fakfak Animal Husbandry Office. The buffaloes, goats, pigs, and horses’ blood samples were collected in 5 ml EDTA tubes from the jugular vein located in the ventrolateral area of the neck using vacutainer needles, size 16–18. Meanwhile, the dog’s blood was drawn from the cephalic antebrachial vein in the leg using a size 21 vacutainer needle. By using a sterile micropipette, approximately 10 ul of EDTA blood was dropped onto a microscope slide, then smeared and stained with Giemsa (MERCK Millipore, Germany) for Plasmodium microscopic identification, while the remaining blood was dropped by sterile micropipet (about 20 ml) onto a filter paper (Whatman CAT No. 1442-090) until it absorbed to about 1.5 cm in diameter and hold back until blood at filter paper dry. The dry filter paper was put on a sterile clip seal plastic bags and stored at room temperature for a maximum of 10 days. All the sample collection process was done by sterile conditions.
A dried blood spot (DBS) isolation kit for DNA extraction on filter paper (Cat. no. 36000) from Norgen Biotec was used. A 6 × 3 mm piece of blood-stained filter paper was put into a 1.5 ml tube containing 100 μl of digestion buffer B. It was vortexed and incubated at 85°C for 10 minutes. Afterwards, 20 μl of proteinase K and 300 μl of lysis buffer B were added to the tube and then vortexed before incubation at 56°C for 10 minutes. About 250 μl of 95% ethanol was added to the tube and then vortexed, while the DNA content was washed by adding 500 μl of WN wash solution and centrifugated for one minute at 8,000 rpm. Washing was carried out again using 500 μl of WN wash solution and centrifugated at 14,000 rpm. For DNA elution, 90 μl of elution buffer B was put into the tube and centrifuged at 8,000 rpm for one minute, and the purified DNA was stored at -20°C.
DNA amplification by nested PCR and qPCR were performed as directed by Tiangen Biotech (Beijing). Plasmodium DNA amplification was carried out using the nested PCR method with a 2× Tag Plus PCR mix enzyme (Tiangen). The final volume of 12.5 μl contained 6.25 μl enzyme, 2.25 μl ddH2O, 1 μl forward primers, 1 μl reverse primers, and 2 μl DNA sample. For sequencing, the PCR mixture’s volume was doubled, with the final volume being 25 μl, while the primer sequences of P. falciparum, P. vivax (Snounou et al., 1993a) and P. knowlesi (Lee et al., 2011) can be seen in Table 1.
The nested one DNA amplification temperature was set at 94°C denaturation (one minute), 55°C annealing (one minute) and 72°C extension (one minute) for 35 cycles. For nested two, denaturation was carried out at 94°C (30 seconds), 55°C annealing (one minute) and extension was at 72°C (30 seconds) in 35 cycles. There was a difference in the annealing temperature for each species in nested two, namely 55°C (one minute) for PCR multiplex P. falciparum and P. vivax, but 56°C (one minute) for P. knowlesi. Nested one products were used as templates for nested two and both were run on agarose gel 1.5% and 2%, respectively, while qPCR was run on agarose gel 1.5% and view in a gel documentation system. All the stage of DNA amplification was carried out in sterile media and places such us laminar airflow. Molecular work was not performed for P. ovale and P. malariae due to difficulties in finding the positive control, and according to the local health office these species have never been reported from Sumba and Fakfak.
Preventing the possibility of positive contamination of DNA Plasmodium in the first extraction, DNA was re-extracted from the same blood spot at filter paper. The extraction room used was confirmed to have never been used for Plasmodium extraction sample previously. Filter papers were cut by scissors which has been sterilization. PCR was performed using the primers, rPF1 and rPF2, as well as rPV1 and rPV2 (Snounou et al., 1993b) to detect P. falciparum and P. vivax, respectively. The same extraction and amplification method were used as described above.
To determine the Plasmodium species, in the second round of nested PCR, products having positive band targets were sent to the 1st BASE, Axil Scientific Pte Ltd Singapore for sequencing. The DNA sequence result was adjusted using multiple alignments found in the BioEdit 7.0 application (Hall et al., 2011) and then read by the BLAST program from the NCBI website.
This study was approved for ethical clearance by the ethics committee of the Faculty of Medicine, Hasanuddin University (734/H4.8.4.5.31/PP36-KOMETIK/2018). All efforts were made to ameliorate any suffering of animals. To prevent stress, animals were comforted by their owners while blood samples were taken, and sampling was performed by experienced officers. Second and third blood samples were taken if there was a failure in the first sample and only if the animals were cooperative. About 20% of animals were sampled more than once.
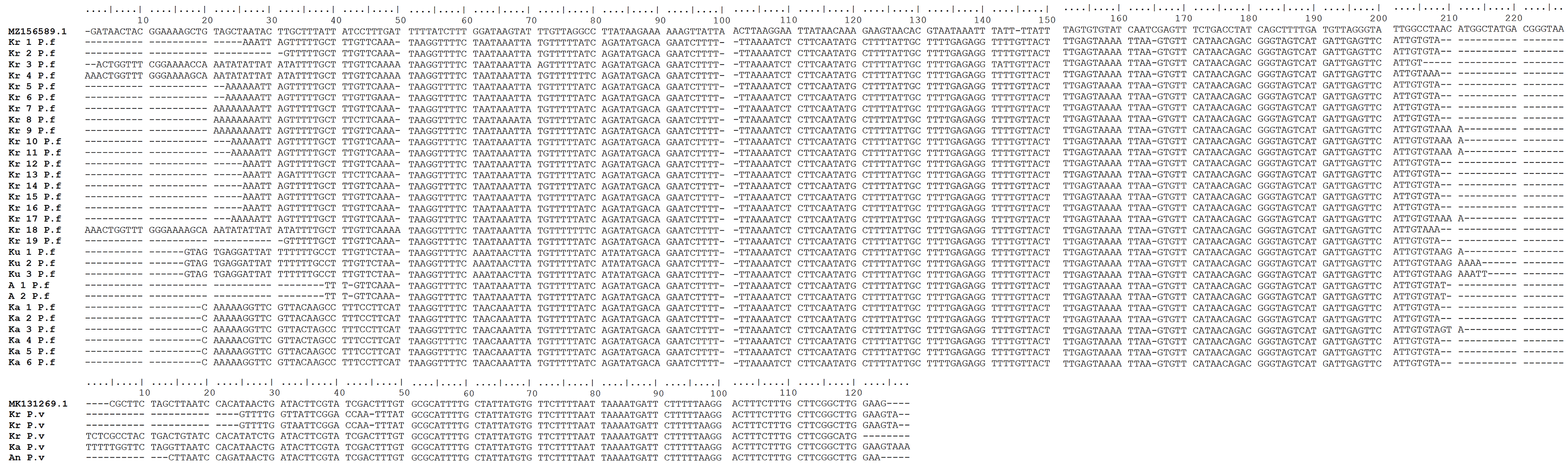
A total of 208 and 62 blood samples were collected in Gaura and Fakfak villages, respectively, from 92 buffalos, 21 horses, 121 goats, 18 dogs, and 18 pigs. Tests conducted using the nested PCR method identified 32 of the 270 animals as positive for P. falciparum and P. vivax. Furthermore, 20.7% buffalo, 14.3% horse, 5.8% goat, and 16.7% dog were Plasmodium-positive, with one buffalo showing mixed infections of P. falciparum and P. vivax. P. knowlesi was absent in all the blood samples and no form of malaria parasites was found in the 18 pigs. Additionally, PCR gel products, DNA sequence results, and the quality of samples can be seen in Figures 2, 3, and 4, respectively (Munirah et al., 2021). Plasmodium distribution in samples from Gaura and Fakfak are presented in Table 2, showing that blood containing the malaria parasites was only found in Gaura. The results of qPCR performed using rPF1–rPF2 and rPV1–rPV2 primers were similar to those of the nested PCR.
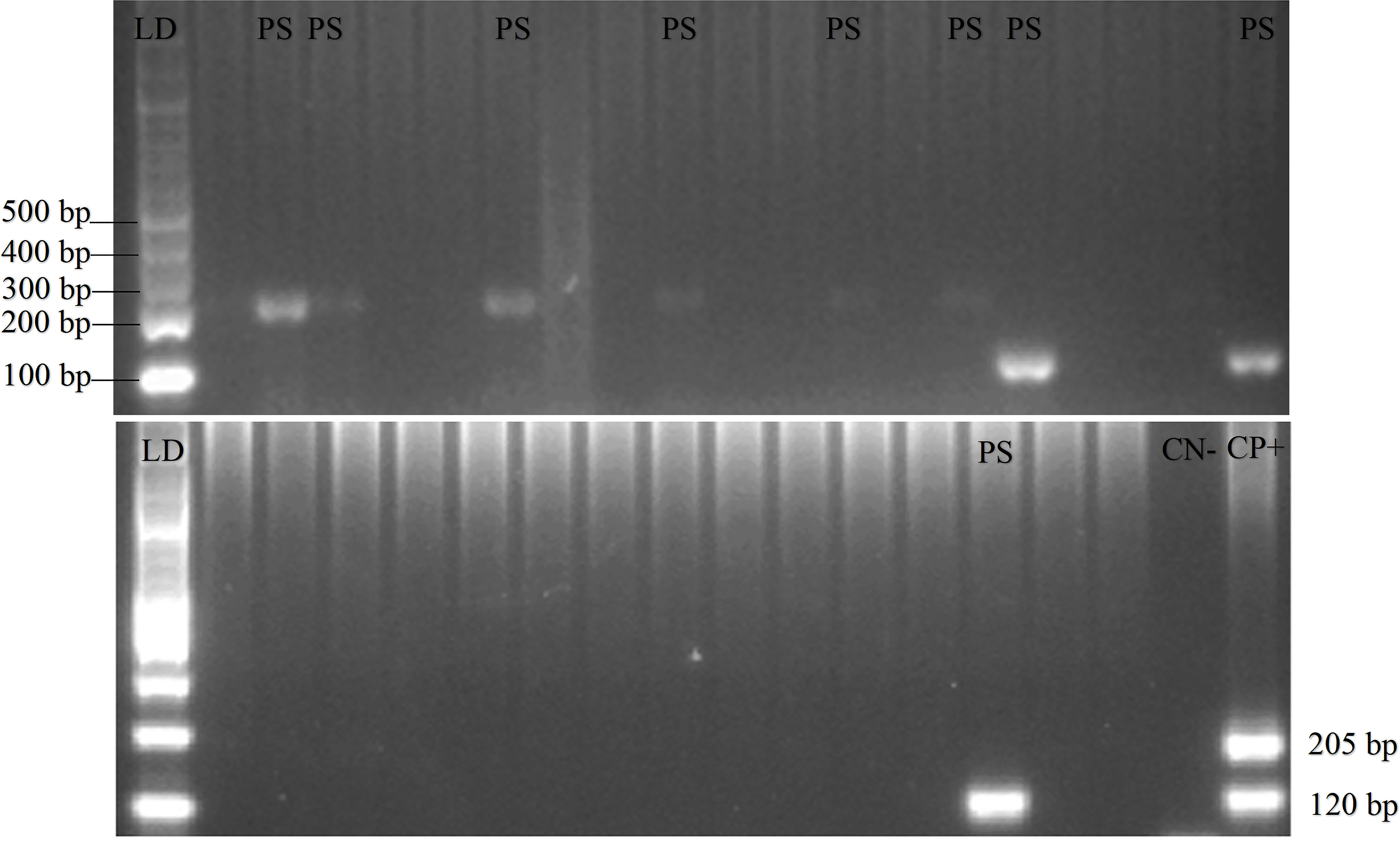
120 bp for positive Plasmodium vivax, 205 bp for positive Plasmodium falciparum.
Microscopically, trophozoites, schizonts, and gametocyte forms at 100× magnification can be seen in Figure 5. P. falciparum gametocytes found in buffaloes were sausage and crescent-shaped (a, b), while schizonts found in horses were smaller or the same size as the red blood cells (c). The P. vivax gametocyte was larger than the red blood cells found in buffalo (d). P. falciparum gametocyte and trophozoites (ring-shaped) with one or two nuclei was found in goats (e) and P. falciparum trophozoite found in horses had one nucleus (f).
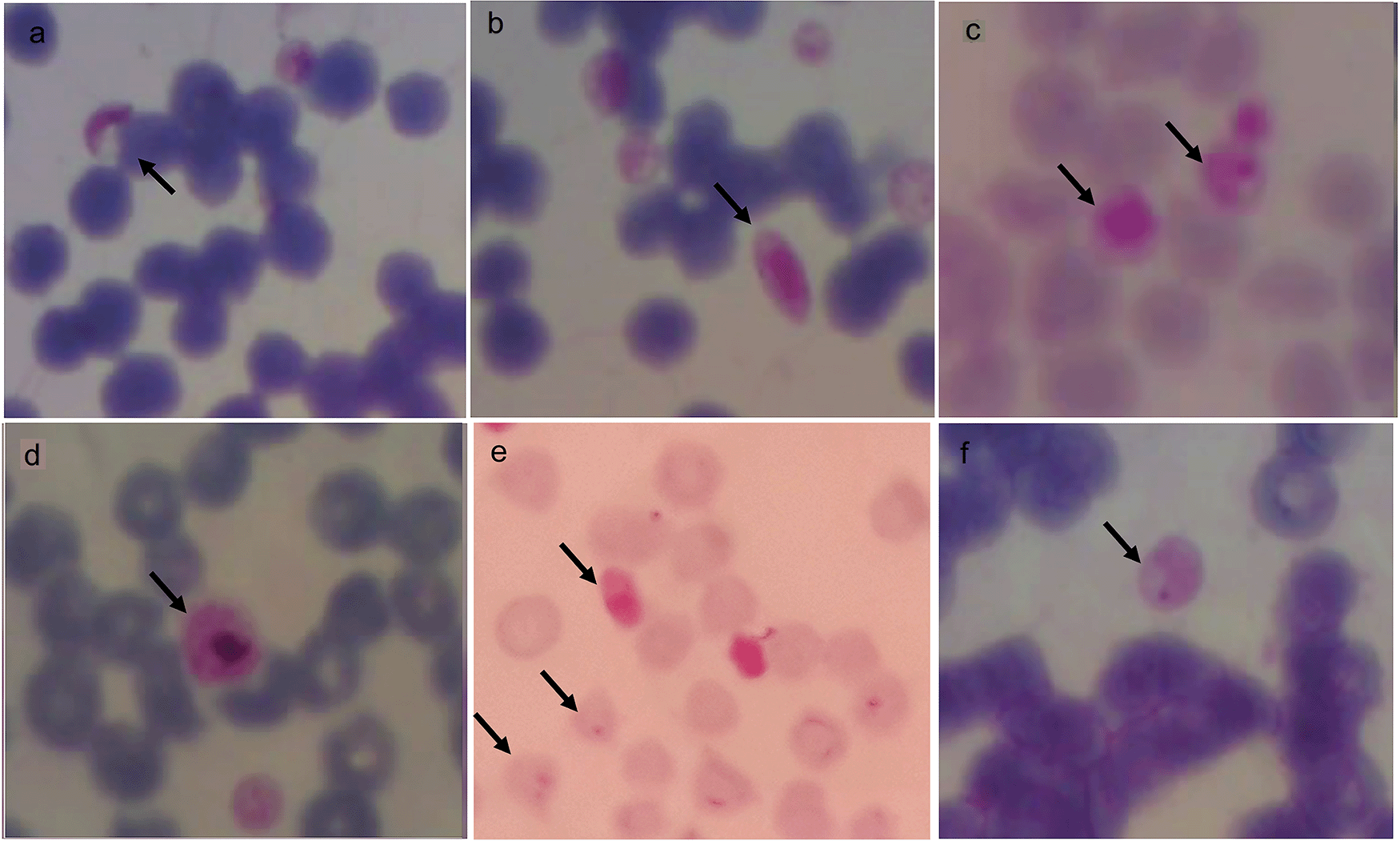
Gametocytes (a,b,d) in buffalo, schizont in horse (c), gametocyte and trophozoite in goat (e) and trophozoite in horse (f) with magnification 1000 ×.
Plasmodium presence was suspected in domestic animals because malaria cases in both Gaura and Fakfak villages remained high despite applying maximum preventive efforts including insecticide-treated bed nets. The results showed that 32 of the 270 blood (11.9%) samples contained human Plasmodium, serving as the first data report regarding this parasite, hence further investigation should be conducted.
Previous studies found P. relictum in avian species (Cox, 2010), P. cephalophi in ungulates (Bruce et al., 1913), P. traguli in mousedeer (Garnham and Edeson, 1962), P. brucei in gray duiker (Bruce et al., 1915; Templeton et al., 2016a), P. bubalis in water buffalo (Sheather, 1919), and P. odocoilei in white-tailed deer (Garnham and Kuttler, 1980; Perkins and Schaer, 2016). Other parasites detected included P. caprae in goats (ruminant) (Kaewthamasorn et al., 2018), P. bergei in Rodentia (Vincke and Lips, 1948), as well as P. cynomolgi, P. inui, and P. fragile in primates (Dixit et al., 2018). The five Plasmodium species infecting humans were originally parasites in primates (Duval et al., 2010; Knowlesi, 1935; Liu et al., 2014; Ng et al., 2008; Singh and Daneshvar, 2013; Zhang et al., 2016). This study identified P. falciparum in buffalos, goats, dogs, and horses, as well as P. vivax in buffalos, goats, and dogs. Initially, Plasmodium presence in the erythrocytes (RBCs) of these animals was uncertain, but the nested PCR produced the same results for all positive samples. Sequencing analysis of positive bands in the nested PCR confirmed the presence of P. falciparum and P. vivax (Figure 3). The number of positive Plasmodium cases in buffalos was higher because of susceptibility to parasitic infections. This was consistent with the report of 10 parasitic infections observed by Mursyid in Central Lombok (Mursyid et al., 2020). Another source declared age as one of the factors predisposing buffalos to high risk of infections (Nurhidayah et al., 2019), with the average age exceeding seven years. This study provides the first report of human Plasmodium in ruminant, ungulate, and carnivorous domestic animals.
Investigations conducted by Templeton in Thailand in 2008 and 2015 showed the presence of P. bubalis among buffalos. The microscopic appearance of P. bubalis was depicted in the journal published by Templeton et al. (2016b). However, this current study did not detect any similarity between P. bubalis and P. falciparum gametocytes in buffalos. A molecular examination method targeting the cytochrome b (cytb) gene identified the presence of P. caprae in goats from Thailand, Myanmar, Iran, Kenya, and Sudan. The trophozoites of P. caprae can be observed in a journal published by Kaewthamasorn et al. (2018).
The discovery of Plasmodium among domestic animals in malaria-endemic areas raises the following questions. How do P. falciparum and P. vivax survive in these animals? Can animals serve as intermediate hosts for these parasites? Have both species evolved to live in ruminants, ungulates, and carnivores? Due to repeated exposure, have these animals become more susceptible to Plasmodium, which generally infects humans? Is Plasmodium pathogenic in animals?
P. knowlesi is identified as a commensal microbe in primates but pathogenic in humans (Jongwutiwes et al., 2004; Ng et al., 2008; Singh and Daneshvar, 2013), and the transmission to humans can be attributed to forest loss or the invasion of primate habitats (Davidson et al., 2019). There is a possibility that the proximity of animals and humans facilitates easier transfer of parasites between both groups through mosquitoes. In humans, P. falciparum and P. vivax infect by initially growing in liver cells before moving to RBCs. The parasites multiply in RBCs, leading to medical conditions characterized by fever, chills, headache, profuse sweating, weakness, rheumatic pain, symptoms of anemia or lack of blood, and nausea or vomiting. Plasmodium found in non-primates remains undetermined as pathogenic or commensal. However, sporozoites of P. brasilianum identified in animals migrate directly to the liver where multiplication occurs, releasing merozoites. The merozoites infect RBCs, initiating symptomatic disease in these animals (Erkenswick et al., 2017). Organisms infected by Plasmodium become symptomatic when the parasite cycle advances to the erythrocyte stage or causes malaria due to the rupture of RBCs. The identification of intermediate hosts for P. falciparum and P. vivax in livestock is still a challenge. Furthermore, various studies attempted to provide evidence for the origination of these two species from chimpanzees and gorillas.
The investigation conducted by Prugnolle in 2013 signified that P. vivax detected in monkeys and humans had similarities. The results showed the possibility of natural transfer between the two organisms, particularly in environments where animals and humans coexist, facilitating continuous parasite transmission through vectors (Prugnolle et al., 2013). Similarly, a study by Mu in 2005 described P. vivax as a zoonotic parasite (Mu et al., 2005).
High API was observed in Fakfak and Gaura villages, but only animals from Gaura had human Plasmodium. This difference may be attributed to the long distance of approximately 50–500 m between the residential houses and animal enclosures in Fakfak. Meanwhile, in Gaura, residents live in stilt houses, with the ground floor and surroundings serving as a shelter for animals, which facilitate microbial transfer between humans and animals through mosquitoes. The inaccessibility of sampling locations and steep geographical conditions in Fakfak posed challenges during sample collections. Moreover, the vector census conducted in both areas identified 11 Anopheles mosquito species in Gaura village, including An. vagus, An. sundaicus, An. aconitus, An. Kochi, An. flavirostris, An. indefinitus, An. maculatus, An. minimum, An. annularis, An. nivipes, and An. subpictus. Molecular examination results showed that two An. sundaicus were infected by Plasmodium, while no Anopheles was found in Fakfak. The species An. sundaicus was identified as zoophilic in India (Vidhya et al., 2019) and anthropophilic in Mekong, Vietnam (Trung et al., 2005). These different reports from both areas suggested variations in the behavior of An. sundaicus, showing the diversity of mosquito biting patterns influenced by environmental factors. The proximity between domestic animals and humans in Gaura village may increase the tendency of bites from An. sundaicus, thereby necessitating further investigations.
The application of livestock for zooprophylaxis in malaria-endemic areas offers several advantages but tends to increase the survival of mosquitoes, which become potential disease vectors. Proximity between infected and uninfected organisms is a significant factor driving parasite transmission (Kuris et al., 1980). The study by Hasyim suggests that livestock in endemic areas have a high potential to increase malaria incidence in Indonesia (Hasyim et al., 2018). Moreover, zoopotentiation often occurs when livestock are kept indoors or near houses (Donnelly et al., 2015). Locating livestock away from humans tends to reduce malaria cases (Franco et al., 2014). Strategies for separating livestock from humans to minimize mosquito bites in both groups are essential. Mosquitoes that have fed on the blood of animals tend to not suck blood from humans. However, closeness between animals and humans can lead to massive pathogen transmission through intermediary vectors. Addressing factors such as vector abundance, mosquito survival, mosquito behavior, and local environment is crucial in mitigating zoonotic potential.
Plasmodium can be detected microscopically due to the smaller size of RBCs in animals compared to humans, but the molecular method is more significant for identifying the presence of this parasite. Nested PCR which offered high sensitivity similar to Real-Time PCR at a low cost was applied for the detection process (Green and Sambrook, 2019; Perandin et al., 2004). The microscopic method using double fluorescent dyes with Giemsa stain is recommended for subsequent studies (Guy et al., 2007).
Further investigation is needed to confirm the presence of domestic animal Plasmodium by amplifying the cytochrome B sequence and sequencing the invasion ligands such as DBL protein. Additionally, other challenges in malaria elimination shown by the results of this study should be addressed.
In conclusion, Plasmodium species previously recognized as human parasites were detected among domestic animals in this study. The obtained results provided insights into malaria elimination challenges in the explored areas and were expected to promote public health improvement while guiding the development of malaria prevention strategies.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)